


微生物腐蚀(MIC)是指微生物及其代谢产物对金属材料的腐蚀破坏过程[1,2]。MIC普遍存在于各种自然环境中,如土壤[3]、油田系统[4,5]和海洋环境[6,7],典型的MIC细菌包括硫酸盐还原菌(SRB)[8]、铁氧化菌(IOB)[9]、硝酸盐还原菌[10]和产酸菌[11]等。其中,由SRB引起的腐蚀损失占MIC的50%以上,SRB被认为是厌氧环境中Fe腐蚀的主要原因之一。SRB通过利用硫酸盐作为末端电子受体,在其生理代谢过程中产生腐蚀性H2S,导致严重的局部腐蚀[12]。最近研究发现,电活性SRB通过胞外电子传递的方式也可以直接从金属表面获取电子,加速金属的腐蚀[13~16]。新型电活性SRB从金属Fe中获得电子的方式比通过H2的方式更直接,许多研究者提出了胞外电子传递(EET)机理解释新型SRB腐蚀过程[17,18]。
电活性微生物能够直接与电极发生反应并对外界电刺激做出响应,电极电位能够影响微生物的活性、丰度和代谢效率[19]。Liduino等[20]研究表明:-1000 mV vs. Ag/AgCl电位下刺激了SRB的氢酶活性,显著促进了点蚀过程,McCully等[21]研究直接证明了电活性IS5细菌能够直接从极化的导电玻璃(ITO)获得电子。Lv等[22]采用FTO电极研究表明,在-1.05 V vs. SCE电位下由于外加电荷、pH值的增加以及电极表面钙质沉积物的形成抑制了SRB的附着,同时SRB的存在也增加了阴极电流。Yu等[16]研究表明SRB生物膜能够从-0.61 V vs. SHE (相当于-0.85 V vs. SCE)极化的石墨电极中获得电子形成H2,析氢反应是SRB生物膜与电极之间发生电子传递的重要过程。
由于在阴极极化表面发生的复杂过程,海洋环境中阴极极化的钢铁电极与SRB之间的相互作用至今尚未得到很好的阐明,同时,极化作用下强腐蚀性SRB的腐蚀机制研究较少,还缺乏钢铁阴极极化电位对SRB电子传递机理的新认知。前期研究已经证明,电活性SRB在-0.85 V vs. SCE电位下能够显著促进细胞电子传递与代谢过程[16],而在-1.05 V电位下能够显著抑制其附着[22],因此,本研究选择这两个电位作为典型阴极极化条件,旨在初步研究强腐蚀性的Desulfovibrio bizertensis SY-1在阴极极化作用条件下对X70管线钢腐蚀行为的作用。本研究将为进一步认识电活性微生物腐蚀机理提供新的认识,也为阴极保护技术在微生物腐蚀防护中的应用提供初步参考。
1 实验方法
X70管线钢试片大小为85 mm × 35 mm × 2 mm (用于腐蚀速率测试)和30 mm × 20 mm × 5 mm (用于腐蚀形貌观察及腐蚀产物分析实验),其化学成分(质量分数,%)为:C 0.130,Si 0.290,Mn 1.650,P 0.020,S 0.010,V 0.034,Nb 0.010,Ti 0.023。实验前用800#、1500#、2000#的碳化硅砂纸打磨,然后用无水乙醇和二次水超声清洗,N2吹干,称重,备用。实验前将试片在紫外灯下照射30 min以除菌。
按照文献[25]方法进行阴极极化实验。采用三电极体系,将灭菌的X70管线钢试片用作工作电极,使用铂片电极夹固定,通过纳米胶密封从而暴露固定表面积;铂片(1 cm × 1 cm)为对电极,饱和甘汞电极为参比电极。分别施加-0.85和-1.05 V vs. SCE的阴极极化电位,以无施加极化电位即处于自然腐蚀电位(标记为OCP)下的试片作为对照,本研究中所指的极化电位均相对于SCE。阴极极化组在30℃恒温条件下培养10 d。
在SRB细胞生长过程中,采用显微镜直接计数法(OLYMPUS DP80)计算细菌的生长曲线[26]。采用离子色谱(Thermo u3000)对培养液中醋酸浓度变化进行测试,采用液相色谱(Thermo DIONEX AQUION RFIC)对培养液中的乳酸浓度变化进行测试。
采用扫描电镜(SEM, FEI Quanta 250)对锈层和生物膜形貌进行观察,加速电压5~20 kV。试片放置于2.5% (v/v)的戊二醛溶液中浸泡4 h,取出后用25%、50%、60%、70%、80%、90%和100%浓度的乙醇(v/v)逐级脱水10 min后喷金。
采用透射电镜(TEM, JEOL 1400plus)进行样品观察,加速电压12 kV。试片用无菌手术刀片刮取6 cm2表面生物膜至无菌的二次水中,使用离心机(4℃, 4000 r/min, 5 min)收集细胞,用无菌去离子水清洗收集的细胞3次,最后,将铜网放在该细胞水溶液中10 min,然后在红外光条件下干燥15 min。
试片用灭菌的PBS缓冲液轻轻冲洗,用活死细胞染色试剂盒(BioVision)染色(活菌为绿色, 死菌为红色),37℃暗处理15 min后在荧光显微镜下(OLYMPUS DP80)用蓝色荧光观察。试片取出后,使用无菌二次水轻轻冲洗,N2吹干,使用激光Raman光谱(HORIBA Jobin Yvon)对腐蚀产物进行分析。Raman光谱采用Nd:YAG激光,激发波长为532 nm。为了尽量减少激光加热对样品的损坏,激光功率始终保持在较低的水平,约为1 mW。
将试片分别浸没在无菌培养基、含有SRB培养基以及分别施加-0.85和-1.05 V vs SCE阴极极化电位的SRB菌液中,10 d后将试片取出,根据ISO 8501:2009,IDT[27],配置酸洗液,清洗试片,以完全去除试片表面的腐蚀产物,然后用去离子水和乙醇清洗,最后用N2干燥,并再次测量每个试片的质量,计算腐蚀前后的质量差,根据下列公式计算试片在SRB培养液中的腐蚀失重:
其中,
通过共聚焦激光扫描显微镜(CLSM, Lext OLS5000)随机选取试片表面15处的腐蚀坑进行数量、直径和深度的测试。
2 结果与讨论
2.1 阴极极化对SRB细胞生长及代谢的影响
为研究不同极化电位(-0.85和-1.05 V)对SRB代谢影响,以自腐蚀环境(OCP)作为对比实验,分别检测OCP体系、-0.85和-1.05 V条件下的代谢因子变化情况。X70钢在SRB培养液中施加不同阴极保护电位的I-t曲线见图1a,施加了-0.85和-1.05 V电位后,试片表面电流会先减小后增大,最终分别稳定在-0.81和-3.41 mA。
图1
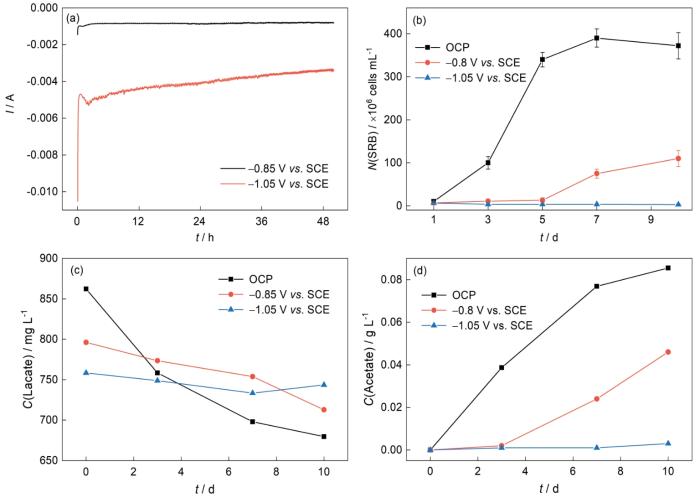
图1
X70钢在SRB培养液中施加不同阴极保护电位的I-t图,浮游细胞计数以及溶液乳酸和醋酸的变化趋势
Fig.1
I-t plot (a), planktonic cell count (b), and the trend of solution lactate (c) and acetate (d) of X70 steel with different cathode potentials applied in SRB culture medium
图1b显示了SRB细胞在自然腐蚀电位、-0.85和-1.05 V电位条件下浮游细胞的生长趋势。在无外加极化电位的培养液中,从第1 d到第7 d,细胞处于对数生长期并迅速繁殖,细胞数量最高达到3.9 × 108 cells·mL-1,7 d后细胞开始下降。施加了-0.85 V的阴极极化后,浮游细胞前5 d数量几乎不变,5 d后细胞开始缓慢增加,直到第10 d达到峰值1.1 × 108 cells·mL-1。在-1.05 V电位下浮游细胞数量在整个培养周期内一直不变。这表明,-0.85 V电位下只能抑制SRB细胞前期5 d的生长,随着培养时间的增加,阴极电位对SRB浮游细菌生长的抑制作用将减弱。
SRB以乳酸为电子供体,硫酸盐为电子受体,其反应如下:
氧化反应:
还原反应:
对SRB在不同条件下培养液中的乳酸和醋酸的变化过程进行测试,如图1c和d所示。在无外加极化的自腐蚀电位下,SRB菌液中乳酸被快速消耗,而在阴极极化电位施加后,其溶液中的乳酸浓度消耗速率明显减缓,这说明阴极极化电位的施加抑制了Desulfovibrio bizertensis SY-1细胞的代谢过程。根据反应(2)和(3),当乳酸被氧化为醋酸时,乳酸的消耗量和醋酸的生长量一致,在-0.85 V电位施加实验中,乳酸的消耗量和醋酸生产量的变化趋势一致,但乳酸的消耗量高于醋酸生产量,这说明有部分的乳酸被代谢为CO2或者其他次级代谢产物。在-1.05 V电位下,浮游SRB细胞几乎没有生长,因此溶液乳酸、醋酸也几乎没有变化。
2.2 生物膜和腐蚀形貌表征
图2显示了X70试片在不同电位下极化10 d后的形貌。对于自然腐蚀电位下的X70试片,其表面被黑色和黄色腐蚀产物覆盖,厚度大约2 mm;在-0.85 V的电位下,试片表面被黄色腐蚀产物覆盖,部分金属基底清晰可见;然而,试片在-1.05 V的电位极化下,几乎没有腐蚀产物,试片表面依然有金属光泽。
图2

图2
X70试片在不同电位条件下的SRB培养液中浸泡10 d后的形貌
Fig.2
Optical morphologies of X70 coupons in the SRB culture medium at OCP (a), -0.85 V (b), and -1.05 V (c) potentials after the 10 d incubation
图3为X70试片在不同电位极化后的表面附着细胞的荧光形貌。在自然腐蚀电位的对照体系中,X70表面存在大量的活细胞(绿点);而在-0.85 V电位极化后的试片表面细胞数量明显减少,并出现了死细胞(红点);在-1.05 V的电位极化下试片表面几乎没有活细胞附着,这说明随着阴极极化电位的施加抑制了试片表面细胞的附着,且随着电位负移,抑制效果更明显。
图3

图3
X70试片在不同电位条件下的SRB培养液中浸泡10 d后表面附着细菌的荧光染色照片
Fig.3
Fluorescence staining photos of attached cells on the surface of X70 steel coupons in the SRB culture medium at OCP (a), -0.85 V (b), and -1.05 V (c) potentials after the 10 d incubation
图4为X70试片表面在不同环境下浸泡10 d后的腐蚀产物和生物膜形态。在无菌培养基中(图4a和b),试片表面形成了均匀且致密的片状腐蚀产物。致密的腐蚀产物层可能会阻碍金属的溶解和溶液离子的扩散,从而对X70管线钢起到保护作用。在自然腐蚀电位下的SRB培养基中(图4c和d),试片表面附着了大量且密集的弧状SRB细胞,其产生的胞外聚合物(EPS)会与附着的细胞、无机腐蚀产物结合在一起,形成致密的生物膜。施加了-0.85 V的极化电位后的SRB体系中(图4e和f),试片表面附着细胞数量显著低于自然腐蚀电位下的试片表面细菌数量,其表面形成的腐蚀产物也较为疏松。在-1.05 V 的极化电位下(图4g和h),试片表面几乎未见附着的细菌细胞,表面腐蚀产物也较为疏松。
图4
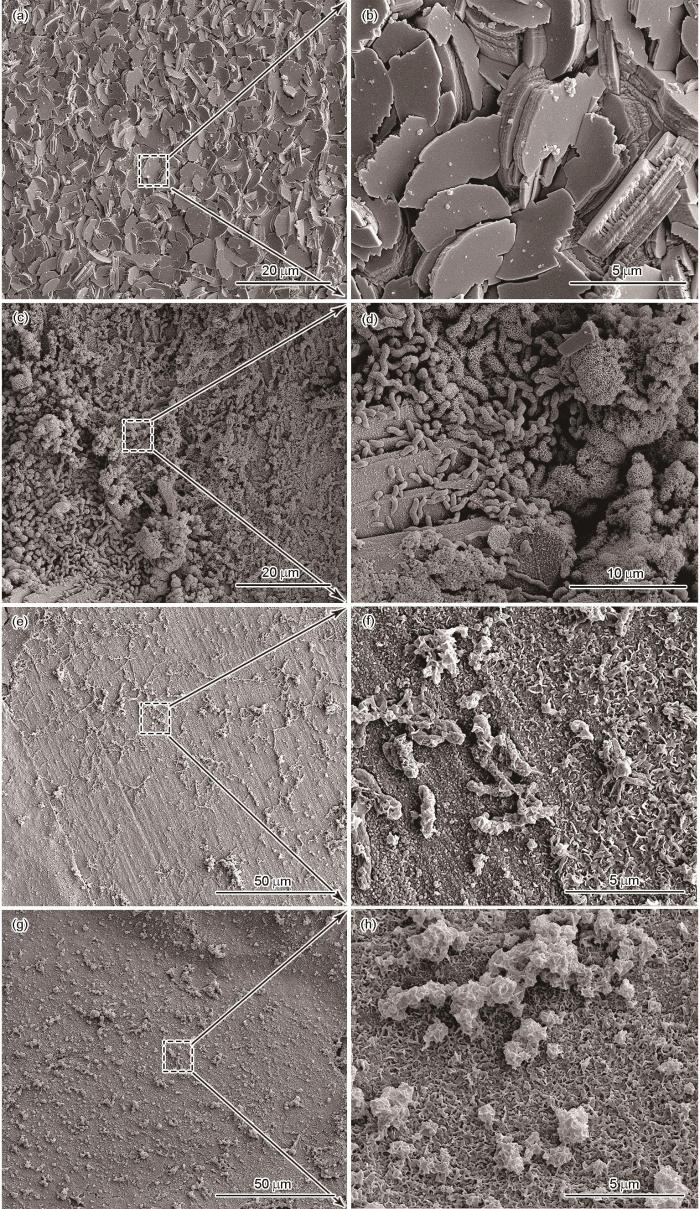
图4
X70试片在无菌和不同电位条件下的SRB培养液中浸泡10 d后的SEM形貌
Fig.4
SEM images of X70 coupons in sterile (a, b) and SRB culture medium at OCP (c, d), -0.85 V (e, f), and -1.05 V (g, h) potentials after the 10 d incubation
利用透射电镜对试片表面附着细胞形态进行观察发现,在不同极化电位下试片表面的SRB细胞形态相似,长度均为1.5~2 µm,有鞭毛。自然腐蚀电位下在X70试片的表面生物膜观察到SRB的细胞代谢产生了球状腐蚀产物,通过腐蚀产物分析发现其主要为铁硫化物。在-0.85 V极化电位下,试片表面发现了大量细胞(图5b),但是其代谢产物FeS与自然腐蚀电位相比明显减少,且SRB细胞鞭毛与相邻SRB细胞相连(图5c),这可能与此极化电位下SRB的代谢方式改变有关;Xu等[27]在杀菌剂为75 mg/L 的四羟甲基硫酸磷(THPS)作用下含有SRB菌液的X70管线钢点蚀坑内也有相似发现,认为可能与胞外电子传递有关。由此推测,在-0.85 V的阴极极化电位和75 mg/L的THPS作用下,SRB对环境的适用和代谢调控有相似的反馈机制。而在-1.05 V的极化电位下,X70管线钢表面观察到的细胞较少,且细胞周围几乎未见SRB细胞的代谢产物。
图5

图5
X70管线钢在OCP、-0.85 V和-1.05 V vs. SCE电位条件下的SRB培养液中浸泡10 d后的透射电镜形貌
Fig.5
TEM images of X70 coupons in the SRB culture medium at OCP (a), -0.85 V (b, c), and -1.05 V (d) potentials after 10 d incubation
2.3 腐蚀产物分析
图6为X70试片在不同条件下腐蚀产物的拉曼光谱分析结果。在无菌培养基中(图6a)浸泡的X70管线钢试片,在243、479、550和953 cm-1处检测到了特征拉曼峰,这是典型的针铁矿(goethite)[28,29],与SEM中观察到的特征的片状形貌对应[28]。在自然腐蚀电位下的SRB培养基中(图6b)的试片上,位置1和2处在283和207 cm-1处有明显的Raman峰,表明位置1和2处主要为典型的纳米四方硫铁矿(mackinawite)[30,31],这种纳米铁硫化物常被认为是非晶态FeS,是从溶解的Fe(II)和S(-II)物种中析出的固体[32,33];此外,位置1和2处的Raman光谱在390 cm-1处有微弱的峰,说明其表面含有少量的针铁矿。对于-0.85 V电位极化下的X70管线钢试片表面,其腐蚀产物组成与自然腐蚀电位下的试片相似,其腐蚀产物主要为四方硫铁矿和少量针铁矿。与前两个体系相比,在-1.05 V的电位极化下试片腐蚀产物的Raman光谱具有较大差异,在302、540和670 cm-1处检测到Raman峰,这是典型的磁铁矿(magnetite)Raman峰[28,34],同时在207 cm-1处也存在较弱的四方硫铁矿峰。
图6

图6
X70试片在无菌和不同电位条件下的SRB培养液中浸泡10 d后的Raman测试结果
Fig.6
Raman analysis of X70 coupons in sterile (a) and SRB culture medium at OCP (b), -0.85 V (c), and -1.05 V (d) potentials after 10 d incubation
2.4 腐蚀失重表征
为了定量分析SRB在阴极极化条件下对X70管线钢的腐蚀影响,对试片在不同极化电位下的SRB菌液中浸泡10 d后进行腐蚀失重测量,结果如图7所示。X70试片在无菌对照体系、自腐蚀电位、-0.85和-1.05 V极化电位下的腐蚀失重为别为0.007 ± 0.002、0.073 ± 0.003、0.034 ± 0.004和0.008 ± 0.001 g/m2。SRB显著促进了X70管线钢的腐蚀(标准差p < 0.05),其腐蚀失重是无菌条件下的10倍。阴极极化能够显著抑制SRB的腐蚀,在-0.85 V的极化电位作用下X70试片的腐蚀失重减少了约50%,-1.05 V的极化电位作用下X70试片的腐蚀失重减少了约91%,仅有在-1.05 V的极化电位下才能有效抑制SRB腐蚀,在该电位下的腐蚀失重与无菌条件下相近。
图7
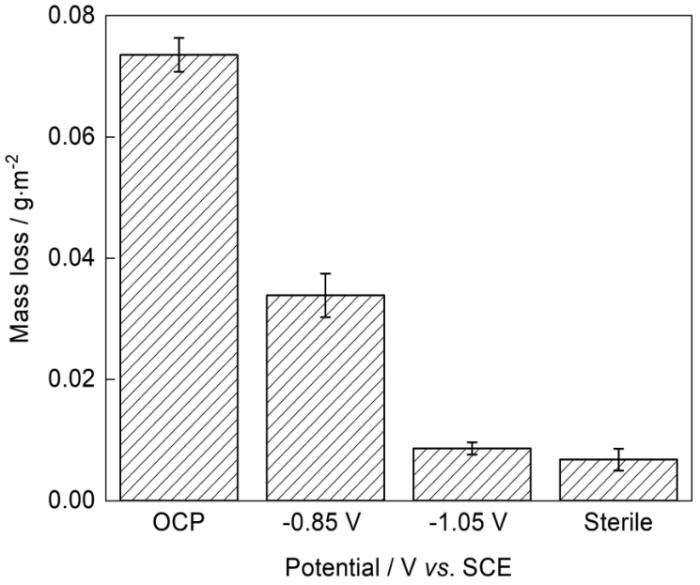
图7
X70管线钢在无菌和不同阴极电位的SRB培养液中浸泡10 d后的腐蚀失重
Fig.7
Mass loss of X70 coupons in sterile and SRB culture mediums with different polarization potential after the 10 d incubation
去除腐蚀产物后,在SRB菌液中试片均可见明显的点蚀,如图8所示。X70试片在无菌对照体系、自腐蚀电位、-0.85 V和-1.05 V极化电位下的平均点蚀坑深度为别为4.2 ± 0.8、22.8 ± 2.4、13.5 ± 1.6和7.7 ± 3.4 μm。自腐蚀电位下的SRB生物膜形成的最大点蚀坑深度为25.4 μm,几乎是无菌条件下最大点蚀坑深度的5倍,SRB显著促进了X70管线钢的点蚀过程,这可能与SRB的胞外电子传递方式有关,即吸附的SRB细胞在EET中可以作为生物阴极,通过直接电子传递(DET)的方式引起严重的点蚀[35~37]。在-0.85和-1.05 V极化电位下的试片点蚀坑深度与自腐蚀电位相比均减少,其最大点蚀坑深度分别为16.6 和10.6 μm。在-0.85 V电位下,其腐蚀速率和点蚀坑深度是无菌条件下的3倍,表明SRB可能会从该电位下获得电子[16],引起试片腐蚀。在-1.05 V电位下X70管线钢的点蚀和腐蚀速率抑制效果最显著,可能在该电位下电极表面的外加电荷、pH的升高以及在电极表面形成的钙质沉积层抑制了SRB的附着[22]。
图8
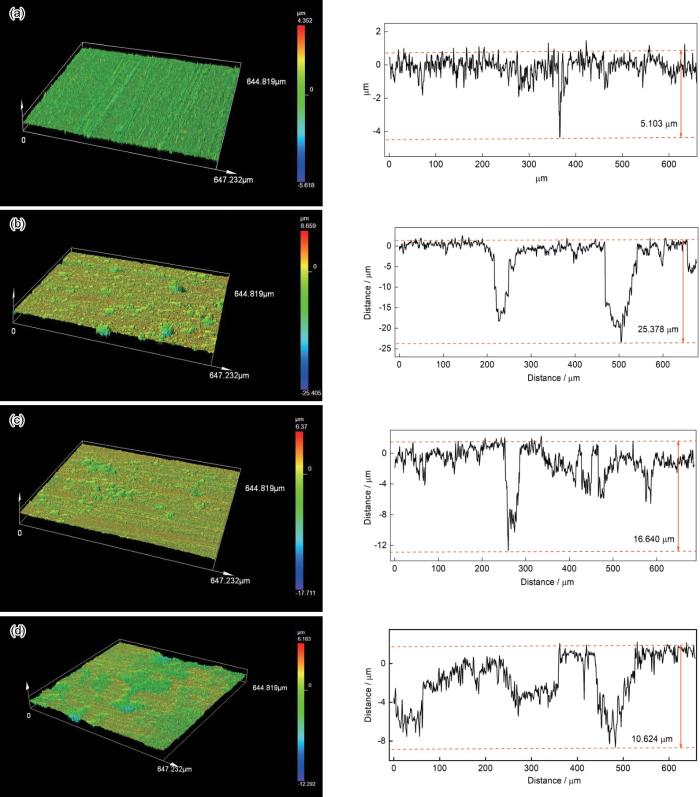
图8
X70试片在无菌和不同电位条件下的SRB培养液中浸泡10 d后的最大点蚀坑深度
Fig.8
Maximum pitting depth of X70 coupons in sterile (a) and SRB culture medium at OCP (b), -0.85 V (c), and -1.05 V (d) potentials after 10 d incubation
3 结论
(1) SRB计数与生理代谢研究表明,在-0.85 V的极化电位下SRB浮游细胞的生长和代谢过程在前5 d受到抑制,随后浮游细胞浓度上升,在-1.05 V电位下SRB浮游细胞浓度受到显著抑制。此外,阴极极化电位下X70管线钢表面附着细胞数量减少,在-1.05 V电位下X70管线钢表面几乎无细胞附着和生长。
(2) 腐蚀产物的拉曼光谱分析表明,在-0.85 V电位与自然腐蚀电位下,试片表面腐蚀产物组成相似,均为四方硫铁矿和针铁矿,而在-1.05 V电位下主要为磁铁矿。
(3) 腐蚀失重分析证明,在-1.05 V电位下失重与无菌条件下基本一致,且最大点蚀坑深度较自然腐蚀电位下减少了75%。
因此,建议在实施阴极保护控制技术之前,应对钢铁所在腐蚀环境中的微生物腐蚀性进行研究和评估,以有利于阴极保护电位的合适选择。
参考文献
Microbiologically influenced corrosion of marine steels within the interaction between steel and biofilms: a brief view
[J].
Marine is the harshest corrosive environment where almost all marine underwater equipment and facilities undergo corrosion caused by marine microorganisms. With the development of marine resources globally, the marine engineering and relevant infrastructures have increased exponentially. Microbiologically influenced corrosion (MIC) leads to severe safety accidents and great economic losses. The specific aggregation of corrosive microbial communities and their interactions with materials conform to a typical ecological adaptation mechanism. On the one hand, corrosive biofilms in the marine environment selectively colonize on a specific steel substrate by utilizing their complex community composition and various extracellular polymeric substances; on the other hand, the elemental composition and surface microstructure of different engineering steels affect the microbial community and corrosive process. MIC in the marine environment is a dynamic process evolving with the formation of corrosive biofilms and corrosion products. In this mini-review, the interactions between corrosive biofilm and steel substrates are explored and discussed, especially those conducted in situ in the marine environment. Herein, the important role of iron in the dynamic process of marine corrosion is highlighted.
Latest research progress of marine microbiological corrosion and bio-fouling, and new approaches of marine anti-corrosion and anti-fouling
[J].Marine resources and industry have become one of the most important pillars in economic development all over the world. However, corrosion of materials is always the most serious problem to the infrastructure and equipment served in marine environment. Researchers have found that microbiologically influenced corrosion (MIC) and marine bio-fouling are two main mechanisms of marine corrosions due to the complicated marine environment and marine organisms. This article summarized the latest research progress about these two mechanisms and indicated that both MIC and marine bio-fouling are closely related to the biofilms on material surfaces formed by the marine microorganisms and their metabolites. As a result, to prevent the occurrence of MIC and bio-fouling, it is important to control the microorganisms in biofilms or prevent the adhesion and formation of biofilms. The traditional method of using chemical bactericide or antifoulant faces the problems of pollution and microorganism resistance. This article introduced four research approaches about the new tendency of applying new materials and technologies to cooperate with traditional chemicals to achieve better and longer effects with lower environment pollution through synergistic actions. Finally, some future research tendencies were proposed for whole marine anti-corrosion and anti-fouling areas.
Microbial corrosion behavior of X70 pipeline steel in an artificial solution for simulation of soil corrosivity at daqing Area
[J].
X70管线钢在大庆土壤环境中微生物腐蚀行为研究
[J].
Microbiologically influenced corrosion of steel in coastal surface seawater contaminated by crude oil
[J].Petroleum-hydrocarbons spilt in surface seawater may pose potential threats to the corrosion of steel infrastructures. We show that crude oil accelerated steel corrosion mainly by accelerating microbiologically influenced corrosion (MIC). Crude oil led to the dominance of marine oil-degraders including Alcanivorax and Marinobacter in both seawater and steel rust, rather than sulfate-reducing bacteria (SRB) which dominated the rust microbial community in the no-oil group. Crude oil not only enhanced microbial oxygen respiration and aerobic hydrocarbon degradation but also nitrate reduction and anaerobic hydrocarbon degradation process in steel rust, indicating more heterogeneous microenvironments formed on steel surfaces. Furthermore, the low abundance of SRB and dissimilatory sulfate reduction gene (dsr), and the existence of iron-carbonate and iron-sulfate minerals implied that microbial sulfide, previously regarded as the main cause of MIC, was not the major contributor to steel corrosion in early petroleum-polluted seawater. Marine specialized oil-degraders seem to play more significant roles under such conditions.
Case analysis of microbial corrosion in product oil pipeline
[J].
成品油输送管道微生物腐蚀案例分析
[J].
The role of biofilms in the corrosion of steel in marine environments
[J].
A critical review of marine biofilms on metallic materials
[J].The formation of multi-species biofilms on marine infrastructure costs the global economy US $ billions annually, resulting in biofouling and microbiologically influenced corrosion. It is well documented that complex biofilms form on almost any submerged surface, yet there are still no truly effective and environmentally friendly treatment or prevention options available. An incomplete fundamental understanding of natural biofilm development remains a key limitation for biofilm control measures. The purpose of this review is to compile the current literature and knowledge gaps surrounding the development of multi-species biofilms in marine conditions on metals.
Research progress on sulfate-reducing bacteria induced corrosion of steels
[J].
硫酸盐还原菌对钢材腐蚀行为的研究进展
[J].
Effect of combined potential polarization on corrosion of X65 steel in seawater inoculated with iron oxiding bacteria
[J].
海水环境中组合电位极化对铁氧化菌腐蚀的影响
[J].采用电化学测试、扫描电镜、激光共聚焦显微镜、Raman光谱等手段研究了-850 mV (vs. SCE,下同) 转-1050 mV和-1050 mV转-850 mV组合电位阴极极化对X65钢在含铁氧化菌 (IOB) 的海水中腐蚀的影响。结果表明:两种组合电位极化都对IOB腐蚀有一定抑制作用;极化与开路电位下X65钢腐蚀产物种类差别不大,含量有区别。-1050 mV极化可以抑制IOB附着但不能完全去除已形成的生物膜,这是-1050 mV转-850 mV极化保护效果优于-850 mV转-1050 mV极化的原因。
Corrosion mechanism of nitrate reducing bacteria on X80 steel correlated to its intermediate metabolite nitrite
[J].
Severe microbiologically influenced corrosion of S32654 super austenitic stainless steel by acid producing bacterium Acidithiobacillus caldus SM-1
[J].
Sulphate-reducing bacteria (SRB) and biocorrosion
[A]. LiengenT, FéronD, BasséguyR, et al. Understanding Biocorrosion[M].
Iron corrosion by novel anaerobic microorganisms
[J].
Corrosion of carbon steel influenced by anaerobic biofilm in natural seawater
[J].
Accelerated anaerobic corrosion of electroactive sulfate-reducing bacteria by electrochemical impedance spectroscopy and chronoamperometry
[J].
Characteristics of hydrogen evolution and oxidation catalyzed by Desulfovibrio caledoniensis biofilm on pyrolytic graphite electrode
[J].
Anaerobic microbiologically influenced corrosion mechanisms interpreted using bioenergetics and bioelectrochemistry: a review
[J].Microbiologically influenced corrosion (MIC) is a major cause of corrosion damages, facility failures, and financial losses, making MIC an important research topic. Due to complex microbiological activities and a lack of deep understanding of the interactions between biofilms and metal surfaces, MIC occurrences and mechanisms are difficult to predict and interpret. Many theories and mechanisms have been proposed to explain MIC. In this review, the mechanisms of MIC are discussed using bioenergetics, microbial respiration types, and biofilm extracellular electron transfer (EET). Two main MIC types, namely EET-MIC and metabolite MIC (M-MIC), are discussed. This brief review provides a state of the art insight into MIC mechanisms and it helps the diagnosis and prediction of occurrences of MIC under anaerobic conditions in the oil and gas industry.
Progress on the corrosion mechanism of sulfate-reducing bacteria in marine environment on metal materials
[J].
海洋环境硫酸盐还原菌对金属材料腐蚀机理的研究进展
[J].硫酸盐还原菌 (SRB) 是一类广泛存在于自然环境中可以利用硫酸盐类物质作为呼吸代谢电子受体的厌氧类微生物,是造成金属腐蚀破坏和设备故障的主要原因之一,已经成为一个重要的研究课题。由于微生物活动的复杂性,生物膜内SRB与金属表面的相互作用缺乏深入的研究,其诱导腐蚀机理和腐蚀过程尚不清楚,难以进行有效的腐蚀预测。基于此,本文从SRB生物膜的呼吸代谢角度介绍了其诱导金属腐蚀的研究进展。介绍了SRB的生态特征和厌氧呼吸过程,重点综述了SRB腐蚀机理,包括阴极去极化、代谢产物腐蚀、浓差电池作用和胞外电子传递等理论,最后简要介绍了微生物腐蚀 (MIC) 研究的方法与技术手段。
Progress on influence of cathodic polarization on sulfate-reducing bacteria induced corrosion
[J].
阴极极化对硫酸盐还原菌腐蚀影响的研究进展
[J].
SRB-mediated corrosion of marine submerged AISI 1020 steel under impressed current cathodic protection
[J].
Direct cathodic electron uptake coupled to sulfate reduction by Desulfovibrio ferrophilus IS5 biofilms
[J].
The effect of cathodic polarization on the corrosion behavior of X65 steel in seawater containing sulfate‐reducing bacteria
[J].
Steel rust layers immersed in the south China sea with a highly corrosive Desulfovibrio strain
[J].Although it is well known that microbes play a significant role in marine corrosion, few studies have systematically studied the relationship between microorganisms and corrosion products under long-term immersion. In this study, the corrosion characteristics of the rust layer formed on carbon steel immersed in the South China Sea for 5.5 years were investigated using various surface analysis and microbial community analysis techniques. Magnetite (Fe3O4), iron sulfide, and green rust were identified in the inner rust layer. The middle rust layer was composed of maghemite (γ-Fe2O3), and some Fe3O4 and mackinamite were also detected. The outer rust layer contained several Fe(III)-oxyhydroxides, and it had a large number of fouling organisms attached to it. In all of the rust layers, anaerobic sulfate-reducing bacteria (SRB) were the dominant bacteria, and they may have played a key role in the formation of the corrosion products. One SRB strain (Desulfovibrio bizertensis SY-1) with a highly corrosivity (13.561 mg/cm2) was isolated from these rust layers, and its physiological and metabolic characteristics were studied. These results expand the membership of corrosive SRB and establish a better understanding of marine microbiologically influenced corrosion (MIC).
Temporal transcriptomic analysis as Desulfovibrio vulgaris hildenborough transitions into stationary phase during electron donor depletion
[J].\n Desulfovibrio vulgaris\n was cultivated in a defined medium, and biomass was sampled for approximately 70 h to characterize the shifts in gene expression as cells transitioned from the exponential to the stationary phase during electron donor depletion. In addition to temporal transcriptomics, total protein, carbohydrate, lactate, acetate, and sulfate levels were measured. The microarray data were examined for statistically significant expression changes, hierarchical cluster analysis, and promoter element prediction and were validated by quantitative PCR. As the cells transitioned from the exponential phase to the stationary phase, a majority of the down-expressed genes were involved in translation and transcription, and this trend continued at the remaining times. There were general increases in relative expression for intracellular trafficking and secretion, ion transport, and coenzyme metabolism as the cells entered the stationary phase. As expected, the DNA replication machinery was down-expressed, and the expression of genes involved in DNA repair increased during the stationary phase. Genes involved in amino acid acquisition, carbohydrate metabolism, energy production, and cell envelope biogenesis did not exhibit uniform transcriptional responses. Interestingly, most phage-related genes were up-expressed at the onset of the stationary phase. This result suggested that nutrient depletion may affect community dynamics and DNA transfer mechanisms of sulfate-reducing bacteria via the phage cycle. The putative\n feoAB\n system (in addition to other presumptive iron metabolism genes) was significantly up-expressed, and this suggested the possible importance of Fe\n 2+\n acquisition under metal-reducing conditions. The expression of a large subset of carbohydrate-related genes was altered, and the total cellular carbohydrate levels declined during the growth phase transition. Interestingly, the\n D. vulgaris\n genome does not contain a putative\n rpoS\n gene, a common attribute of the δ-\n Proteobacteria\n genomes sequenced to date, and the transcription profiles of other putative\n rpo\n genes were not significantly altered. Our results indicated that in addition to expected changes (e.g., energy conversion, protein turnover, translation, transcription, and DNA replication and repair), genes related to phage, stress response, carbohydrate flux, the outer envelope, and iron homeostasis played important roles as\n D. vulgaris\n cells experienced electron donor depletion.\n
Influence of sulfate-reducing bacteria on the corrosion behavior of high strength steel EQ70 under cathodic polarization
[J].
Effects of ferrous ion concentration on microbiologically influenced corrosion of carbon steel by sulfate reducing bacterium Desulfovibrio vulgaris
[J].Ferrous ion (Fe2+) in a sulfate reducing bacteria (SRB) culture medium is known to enhance microbiologically influenced corrosion (MIC) of carbon steel, but the underlining mechanism is controversial. This work showed that it was due to better sessile cell growth that was likely attributed to Fe2+ detoxification of H2S. Two hundred ppm (w/w) initial Fe2+ in the ATCC 1249 medium achieved a 4.7 times higher Desulfovibrio vulgaris sessile cell count and 5.0 times higher C1018 carbon steel weight loss, compared to 20 ppm. Linear polarization resistance, electrochemical impedance spectroscopy and potentiodynamic polarization measurements corroborated weight loss and pitting data trends.
Inadequate dosing of THPS treatment increases microbially influenced corrosion of pipeline steel by inducing biofilm growth of Desulfovibrio hontreensis SY-21
[J].
SEM/micro-Raman characterization of the morphologies of marine atmospheric corrosion products formed on mild steel
[J].
Raman microspectroscopy of some iron oxides and oxyhydroxides
[J].
Corrosion of carbon steel in marine environments: role of the corrosion product layer
[J].
Characterisation of mackinawite by Raman spectroscopy: effects of crystallisation, drying and oxidation
[J].
Characterization of synthetic nanocrystalline mackinawite: crystal structure, particle size, and specific surface area
[J].
High resolution transmission electron microscopic study of synthetic nanocrystalline mackinawite
[J].
Corrosion behaviour of a quenched and partitioned medium carbon steel in 3.5 wt.% NaCl solution
[J].
Long-term survival of Desulfovibrio vulgaris on carbon steel and associated pitting corrosion
[J].
A study on bacteria-assisted cracking of X80 pipeline steel in soil environment
[J].
Toward a better understanding of microbiologically influenced corrosion caused by sulfate reducing bacteria
[J].Sulfate reducing bacteria (SRB) are often the culprits of microbiologically influenced corrosion (MIC) in anoxic environments because sulfate is a ubiquitous oxidant. MIC of carbon steel caused by SRB is the most intensively investigated topic in MIC because of its practical importance. It is also because biogenic sulfides complicate mechanistic SRB MIC studies, making SRB MIC of carbon steel is a long-lasting topic that has generated considerable confusions. It is expedient to think that biogenic H2S secreted by SRB acidifies the broth because it is an acid gas. However, this is not true because endogenous H2S gets its H+ from organic carbon oxidation and the fluid itself in the first place rather than an external source. Many people believe that biogenic H2S is responsible for SRB MIC of carbon steel. However, in recent years, well designed mechanistic studies provided evidence that contradicts this misconception. Experimental data have shown that cathodic electron harvest by an SRB biofilm from elemental iron via extracellular electron transfer (EET) for energy production by SRB is the primary cause. It has been demonstrated that when a mature SRB biofilm is subjected to carbon source starvation, it switches to elemental iron as an electron source and becomes more corrosive. It is anticipated that manipulations of EET related genes will provide genetic-level evidence to support the biocathode theory in the future. This kind of new advances will likely lead to new gene probes or transcriptomics tools for detecting corrosive SRB strains that possess high EET capabilities.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}