
于利宝 , 王彬彬
, 王彬彬
YU Libao, WANG Binbin
中图分类号: TG172.4
文章编号: 1005-4537(2018)01-0010-08
通讯作者:
收稿日期: 2017-01-14
网络出版日期: 2018-02-15
版权声明: 2018 《中国腐蚀与防护学报》编辑部 《中国腐蚀与防护学报》编辑部
基金资助:
作者简介:
作者简介 于利宝,男,1990年生,硕士生
展开
摘要
采用电化学阻抗谱 (EIS)、极化电位扫描等电化学技术和微观形貌观察方法研究含硫酸盐还原菌 (SRB) 的酸性红壤环境中Q235钢的微生物腐蚀 (MIC) 行为及对应电化学过程特征。结果表明:酸性红壤环境中,前4 d为环境适应期,期间SRB细菌数量减少,SRB对腐蚀电化学过程没有显著影响;生长期中SRB促使Q235钢的自腐蚀电位和极化电阻降低,腐蚀速率增大;EIS极化电阻测试结果表明,有菌红壤中腐蚀速率约为无菌红壤中的2倍。SRB呼吸代谢活动可与红壤颗粒表层FeOOH等铁氧化物作用,引起FeOOH的微生物异化还原,促进Q235钢的腐蚀电化学过程。
关键词:
Abstract
Microbial corrosion induced by sulphate-reducing bacteria (SRB) for carbon steel Q235 beneath coating defects was studied by means of electrochemical impedance spectroscopy (EIS), polarization measurement and microscopic surface observation. Results showed that, in acid red soil environment, SRB have no significant effect on the electrochemical process during the initial environmental adaptation period. Then in the next period, the respiratory metabolic activities of the growing SRB lead to decrease of the free corrosion potential of Q235 steel and accelerate corrosion process of the carbon steel. Bacteria can react with iron oxides in the red soil, causing microbial dissimilatory reduction of iron oxides, which promotes electrochemical corrosion process of the carbon steel.
Keywords:
硫酸盐还原菌 (SRB) 广泛存在于土壤环境中,是影响管道、油气井等地下金属设施微生物腐蚀 (MIC) 的主要厌氧菌[1]。SRB通过分泌胞外聚合物形成生物膜粘附于金属表面,加速材料的腐蚀。SRB一般利用分子氢、脂肪酸、脂肪烃等有机物作为碳源和电子供体维持其生命所必需的能量。微生物一般引发金属局部点蚀。据报道,存在SRB的土壤环境中钢铁的腐蚀速率高达0.7~7.4 mm/a[2]。尽管人们对SRB作用下钢铁腐蚀行为已有较为深入认识,但由于微生物过程的复杂性,目前尚无统一的腐蚀机理模型,其中引用较多的有阴极去极化[3]、代谢产物腐蚀[4]、阳极催化和直接电子转移[5]等理论。大量案例分析表明,MIC过程通常涉及多种细菌及代谢产物,很难由单一机制进行描述和解释。
一般认为,SRB的呼吸过程为硫酸盐呼吸,SRB以SO42-为电子受体氧化有机物,通过对有机物的异化作用,获得能量维持生命活动。有研究表明,SRB呼吸途径多样,硫酸盐不是其生存和生长的必要条件,在SO42-缺乏环境中,SRB可很快调整呼吸代谢行为,通过其它的呼吸代谢方式生存和生长,如利用Fe(III),Mn(IV) 等作为末端电子受体[6,7]。这种微生物能量代谢方式普遍存在于沉积物和土壤环境中,其特点是包含电子在微生物细胞和细胞外电子受体/供体之间的传递。
酸性红壤广泛分布于华南热带和亚热带地区,是我国典型的强腐蚀性土壤之一。研究表明,红壤的高腐蚀性与红壤特定的黏土矿物有关,红粘土矿物颗粒表面以赤铁矿 (α-FeOOH) 和针铁矿 (α-Fe2O3) 形式富集的Fe氧化物,在腐蚀过程中作为阴极去极化剂,促进钢的腐蚀[8,9,10]。微生物也是促成红壤腐蚀的重要因素之一,在现场埋设实验中多有提及[11],但对其中涉及的具体过程及机制尚未见深入研究。考虑到红壤黏土矿物颗粒及微生物特性,作者认为,在红壤环境中SRB可能通过胞外呼吸与土壤或腐蚀产物中的FeOOH/Fe2O3等发生作用,影响碳钢的腐蚀动力学过程。
本文采用电化学阻抗谱 (EIS)、极化曲线、循环伏安 (CV)曲线等电化学技术,结合扫描电镜(SEM)、激光共聚焦显微镜 (CLSM) 对表面形貌进行分析,研究红壤中SRB与铁氧化物共同作用下Q235钢的电化学腐蚀行为,试图通过研究金属-介质-微生物界面现象,更深入认识土壤微生物腐蚀行为及机理。
实验所用土壤取自国家材料环境腐蚀试验站江西鹰潭酸性土壤中心站。土壤取自地下1 m,经自然风干,机械碾磨后使其通过孔径为1 mm的土壤筛,经高温灭菌后备用。鹰潭土壤为第四纪红粘土,pH值为4.2,其理化性质详见文献[8]。
实验材料选用Q235钢,其化学成分 (质量分数,%) 为:C 0.30,P 0.019,S 0.020,Si 0.01,Mn 0.42,Fe余量。试样用环氧树脂封装,预留10 mm×10 mm工作面积,模拟涂层破损点。试样工作面用水砂纸逐级打磨至1000#,用去离子水和无水乙醇冲洗,吹干备用。
SRB取自沈阳土壤腐蚀试验站,由API-RP38培养基富集培养。该培养基成分为:乳酸钠4.0 g,酵母膏1.0 g,MgSO47H2O 0.2 g,NaCl 10.0 g,K2HPO4 0.5 g,抗坏血酸0.1 g,(NH4)2Fe(SO4)2 0.02 g,蒸馏水1000 mL,用1 mol/L的NaOH溶液调节pH值至7.0~7.2之间,通高纯N2 2 h以除去培养基中的O2,高温灭菌后低温保存。菌种保存在4 ℃环境中,使用前在恒温箱 (30 ℃) 活化12 h。
取700 g灭菌干燥红壤与300 g灭菌去离子水混成泥浆,接菌组将15 mL含有细菌的培养基接种到1000 g红壤泥浆中,灭菌对照组实验中用15 mL灭菌培养基代替细菌培养基。实验前,两种泥浆试验装置均经通N2除O2后密封。实验在30 ℃恒温水浴锅中进行,实验周期20 d。通过内置的橡胶管,定期抽取接菌体系的泥浆,采用MPN计数法测量SRB的数量。
采用经典的三电极测试系统,工作电极为Q235钢,辅助电极为大面积铂片,参比电极为饱和甘汞电极 (SCE)。电化学阻抗谱测量采用PRSTAT 2273测试系统,激励信号为10 mV的正弦交流波,测试频率范围为105~10-2 Hz,数据由ZSimpWin软件拟合处理。微生物 (膜) 的电活性特征由其在玻碳电极上的循环伏安扫描表征。循环伏安 (CV) 曲线和Tafel极化曲线电位扫描速率分别采用5和0.5 mV/s。
实验结束后,将试样浸入除锈剂 (500 mL盐酸+500 mL去离子水+20 g六次甲基四胺) 中去除腐蚀产物,并用去离子水清洗吹干,使用Philips FEG XL30扫描电子显微镜 (SEM) 和ZEISS LSM 510激光共聚焦显微镜 (CLSM) 观察腐蚀形貌。
测得去离子水和红壤混合后的泥浆pH值为4.1,接入培养基和菌液的泥浆pH值分别为4.5和4.2,中性培养基的加入使泥浆pH值略有上升;菌液由于含有H2S而呈弱酸性,加入菌液后pH值上升幅度较小。实验结束后无菌和接菌环境的pH值分别为4.7和5.1,接菌泥浆的pH值升高较大,可能是SRB阴极氢去极化导致环境中H+减少所致。
图1为接菌泥浆中细菌数量随时间的变化。可见,前4 d细菌数量呈下降趋势,起始数量约为1500 cellmL-1,第4 d降低到约500 cellmL-1。此后呈现指数式增殖,在约9 d达到最大值,约为16000 cellmL-1。此后,随着营养物质的消耗,细菌数量开始快速下降,到20 d细菌数量降至600 cellmL-1以下。SRB适宜生存环境的pH值为近中性,当将SRB接种到pH值为4.2的红壤泥浆中时,有大部分细菌因不能适应这一酸性环境而死亡,导致前4 d细菌数量下降明显[12],存活的SRB适应环境后进入指数生长期。
Q235钢试样在灭菌红壤泥浆中埋设20 d后,试样表面未见明显腐蚀产物,表面有一层红壤颗粒粘附,电极表面已失去原有金属光泽,SEM下可见试样沿划痕发生均匀腐蚀 (图2a和b)。接菌红壤中试样表面也粘附有一层红壤颗粒,除去腐蚀产物后,电极表面呈现严重的溃疡状均匀腐蚀 (图2c和d)。在湿度较高的土壤中,试样表面可形成连续液膜,金属/土壤界面各处电化学差异小,Q235钢发生均匀腐蚀。在接菌环境中试样可见选择性腐蚀,这与钢的微观组织结构有关。这些复杂显微结构造成界面微区相电化学不均匀性,Q235钢表面腐蚀活性点处电子更易于被SRB捕获,利于其获得生长能量。渗碳体 (Fe3C) 是钢中较为稳定的组织,而其周围的铁素体较为活泼,作为活性点而优先溶解,渗碳体则残留在试样表面。CLSM结果 (图3) 显示,接菌红壤中试样的最大点蚀坑深度为14.9 μm,而灭菌环境中试样表面未见明显点蚀坑。
图2 Q235钢在灭菌和接菌水饱和红壤中暴露20 d后的SEM形貌
Fig.2 SEM surface morphologies of Q235 steel after embedding in sterile (a, b) and SRB-inoculated (c, d) red soil for 20 d
图3 Q235钢在灭菌和接菌水饱和红壤中暴露20 d后表面的激光共聚焦形貌比较
Fig.3 CLSM surface morphologies of Q235 steel after embedding in sterile (a) and SRB-inoculated (b) red soil for 20 d
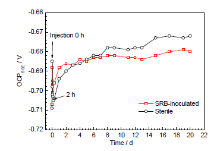
图4为Q235钢在接菌和灭菌两种红壤环境中开路电位 (OCP) 的变化曲线。接菌土壤中试样OCP接菌前为-710 mV,接菌后迅速正移约20 mV,随后2 h内电位下降至-705 mV。灭菌土壤中,OCP在50 h内迅速上升,随后趋于-670 mV附近。接菌环境中试样在前120 h内电位上升明显,最后稳定在-682 mV。
图4还显示,接菌过程中Q235钢的OCP短时间大幅正移,这可能是由培养基中的有机质与金属阳离子在金属表面形成络合物导致[4]。Webster等[13]也认为培养基会造成开路电位正移,对体系造成暂时的保护作用。SRB生物膜形成后,其负电性特征[14,15]导致碳钢的OCP降低。
图4 灭菌和接菌红壤中Q235钢开路电位随时间的变化曲线
Fig.4 Open circuit potentials of Q235 steel in the SRB-inoculated and sterile red soil
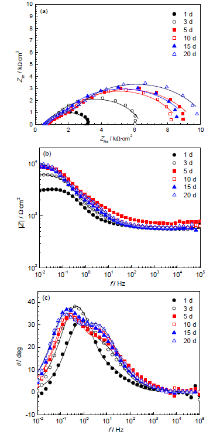
Q235钢在灭菌及接菌红壤泥浆中不同时间的电化学阻抗谱分别示于图5和6。Nyquist图存在两个不完整的扁平容抗弧,高频端容抗弧较小。随时间延长,低频区容抗弧增大 (图5a),阻抗值|Z |逐渐增大 (图5b),相应电荷转移电阻Rct增大。由图5c可见,EIS表现为两个时间常数,在低频区,随容抗弧增大,最大相位角逐渐增大,最大相位角对应的频率逐渐减小,Bode曲线向低频区移动,这是由试样表面逐渐积累的腐蚀产物膜所引起。
图5 Q235钢在灭菌红壤中包埋不同时间的阻抗谱
Fig.5 Nyquist (a), impedance module (b) and phase angle (c) plots of Q235 steel in the sterile red soil
图6 Q235钢在接菌红壤中包埋不同时间的阻抗谱
Fig.6 Nyquist (a), impedance module (b) and phase angle (c) plots of Q235 steel in the SRB-inoculated red soil
实验前期,两种环境中的EIS变化趋势一致,由于培养基和腐蚀产物膜等的影响,|Z |逐渐增大。第5 d,SRB适应环境,开始大量繁殖后,阻抗值降低;生物膜形成后,Bode图左移趋势更加明显;由图5b可以看出,低频区Rct始终比灭菌环境中的低;EIS呈现2个时间常数,最大相位角对应的频率向低频区移动,接菌环境中最大相位角低于灭菌环境中的。
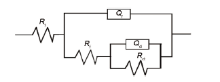
EIS数据均采用图7所示等效电路Rs(Qf(Rf(Qdl-Rct))) 进行拟合。其中,Rs为溶液电阻,Rf和Qf分别是生物膜/腐蚀产物膜电阻和膜电容,Qdl是双电层电容。Qf和Qdl为常相位元件,其阻抗值可表达如下[16]:
其中,Y0表示Q的大小;ω为角频率;n为弥散系数 (0<n<1),反映弥散效应强弱,n越小说明弥散效应越明显,n越接近1表明体系越接近理想电容。
图7 EIS数据拟合的等效电路模型
Fig.7 Equivalent circuit model used for fitting EIS data: Rs(Qf(Rf(QdlRct)))
利用图7所示等效电路拟合EIS,结果示于表1。可见,Rs随包埋时间呈明显减小趋势,红壤电导率低,培养基的加入使环境中可溶性离子增多,随腐蚀反应的进行Rs逐渐降低。Rf随埋设时间的延长呈增加趋势,说明试样腐蚀产物膜随时间不断增厚。此外,Yf在实验过程中保持稳定,而nf呈现降低趋势,说明膜层的电容性质在降低,这可能是随着腐蚀产物膜越来越厚,产物膜层更多地表现出对离子的阻挡作用,而其容纳或接受电子的能力变得不太明显。
表1 灭菌和接菌土红壤中EIS拟合结果
Table 1 Fitting results of EIS in sterile and SRB-inoculated red soil
| Time / d | Rs / Ωcm2 | Yf / Ssncm-2 | nf | Rf / kΩcm2 | Ydl / Ssncm-2 | ndl | Rct / kΩcm2 |
|---|---|---|---|---|---|---|---|
| Sterile soil | |||||||
| 1 | 588.8 | 7.123×10-5 | 0.8335 | 0.3075 | 1.517×10-4 | 0.8770 | 2.366 |
| 3 | 561.1 | 8.340×10-5 | 0.7542 | 1.070 | 1.237×10-4 | 0.9127 | 4.808 |
| 5 | 624.8 | 7.274×10-5 | 0.6603 | 2.538 | 1.304×10-4 | 0.9379 | 6.498 |
| 10 | 605.8 | 8.173×10-5 | 0.7129 | 2.588 | 1.824×10-4 | 0.9661 | 5.579 |
| 15 | 608.9 | 9.339×10-5 | 0.6726 | 3.225 | 2.028×10-4 | 0.9763 | 6.311 |
| 20 | 609.9 | 8.671×10-5 | 0.7065 | 3.353 | 2.403×10-4 | 0.9830 | 6.547 |
| SRB inoculated soil | |||||||
| 1 | 403.1 | 7.729×10-5 | 0.8359 | 0.2622 | 1.465×10-4 | 0.8583 | 2.425 |
| 3 | 401.5 | 8.382×10-5 | 0.7572 | 0.9095 | 1.116×10-4 | 0.8920 | 3.650 |
| 5 | 691.9 | 1.289×10-4 | 0.6731 | 1.128 | 3.962×10-4 | 0.6285 | 3.764 |
| 10 | 693.8 | 1.635×10-4 | 0.6318 | 1.625 | 3.752×10-4 | 0.7712 | 3.376 |
| 15 | 709.9 | 1.796×10-4 | 0.6108 | 2.284 | 3.526×10-4 | 0.9264 | 3.445 |
| 20 | 717.8 | 1.743×10-4 | 0.5998 | 2.635 | 3.425×10-4 | 0.9493 | 3.461 |
接菌环境中试样/红壤泥浆界面的电化学过程受腐蚀产物、活性生物膜和代谢产物的影响。实验前期Rs下降,这主要是受菌液中培养基的影响。Rf随时间也呈增大趋势,但是与灭菌环境相比增加相对较少,SRB经过环境适应期,开始繁殖生长,在钢表面形成生物膜和腐蚀产物膜,导致Qf和Rf升高。
本实验过程中,Q235钢的EIS受金属/土壤界面双电层和生物膜/腐蚀产物膜状态的影响,Rp可表示为:
式中,ZF是体系的Faraday阻抗,即除去溶液电阻Rs和非Faraday阻抗后的阻抗。
据Stern-Geary方程,腐蚀速率可采用Rp-1定性评价[17]。实验过程中,Q235钢的Rp-1的变化示于图8。可见,两体系下Q235钢的腐蚀速率变化趋势相似,但在接菌红壤中的Rp-1明显高于在灭菌红壤中的。起始阶段腐蚀速率较高,实验初期SRB的作用不明显,这与细菌的生长曲线 (图1) 及开路电位变化规律吻合 (图3)。4 d后,SRB适应了酸性土壤环境,开始大量繁殖,腐蚀速率逐渐达到平衡。在灭菌环境中,由于腐蚀产物膜不断积累,腐蚀速率下降,5 d后达到稳定值。此时,在有菌土壤中的腐蚀速率约为无菌土壤中的2倍。Q235钢的腐蚀速率在接菌红壤中总是高于灭菌红壤中的,说明SRB促进了Q235钢的腐蚀过程。
图8 灭菌和接菌红壤中Q235钢极化电阻Rp和Rp-1随时间的变化规律
Fig.8 Polarization resistance (a) and reciprocal of polarization resistance Rp-1 (b) of Q235 steel in red soil as a function of time
循环伏安法是研究电活性生物膜的常用手段,玻碳电极在灭菌和接菌红壤中埋设10 d后测得的循环伏安曲线见图9。可见,接菌红壤中在-1.1和-0.6 V附近出现阴极还原峰1和阳极氧化峰2。在-1.1 V处的阴极还原反应1对应红壤中铁氧化物的还原,而-0.6 V处的氧化峰为Fe(II) 向Fe(III) 的转化;灭菌红壤中,氧化峰减小,阴极还原峰也不明显。
图9 玻碳电极在灭菌和接菌红壤中包埋10 d后的循环伏安曲线
Fig.9 Cyclic voltammetries of a glassy carbon electrode after 10 d embedding in sterile and SRB-inoculated red soil (the scan rate: 5 mVs-1, the electrode area: 0.071 cm2)
由Q235钢试样在红壤中的极化曲线 (图10) 可以看出,两种土壤环境中Q235钢试样均处于活化状态,灭菌红壤中的极化曲线整体居于接菌红壤中的左侧。通过Tafel直线外推法[18]对极化曲线结果进行拟合,Q235钢在灭菌和接菌红壤中的自腐蚀电流密度Icorr分别为5.63和10.0 μAcm-2。
图10 Q235钢在灭菌和接菌红壤中包埋10 d后的极化曲线
Fig.10 Polarization plots of Q235 steel after 10 d embedding in the red soil
Q235钢的土壤环境腐蚀是电化学过程,受土壤中可能存在的阴极去极化剂 (氧化性物质) 控制。铁氧化物是红壤粘土矿物的重要组成部分,其常见存在形式为α-FeOOH (针铁矿) 和α-Fe2O3 (赤铁矿),具有较强的氧化性而容易被还原[8,9,19]。
土壤环境中,SRB可在Q235钢试样表面生成电活性生物膜,并在无外加介体的情况下与基体进行电子交换[20]。微生物会选择电势最低且可用的电子供体使能量增益最大化,以进行新陈代谢;同样,微生物也会选择电势最高且可用的电子受体来获取能量增益,土壤中铁氧化物一般通过这种方式被微生物还原[21]。因此,试样表面微生物膜上存在以乳酸根、H2为电子供体,以铁氧化物为电子受体的反应 (铁呼吸)。土壤中微生物铁呼吸,即以胞外不溶性铁氧化物作为末端电子受体的Fe(III) 异化还原作用,可能是微生物普遍代谢形式之一。从热力学角度分析,Fe(III) 还原比SO42-还原和甲烷化获得更多的能量[7],在低浓度硫酸盐且富含Fe(III) 的环境中,微生物异化还原Fe(III) 应优先于SO42-还原过程[22,23]。
SRB能随环境变化调整自身结构和代谢行为,在SO42-、有机质贫瘠环境中,SRB可利用H2、金属等作为电子供体,以铁氧化物作为末端电子受体[24],实现电子从金属至铁氧化物的转移,在此过程中细菌作为电子传输媒介、铁氧化物作为阴极去极化剂加速材料腐蚀[2,3]。在有机物缺乏环境中,缺少可溶态易利用的电子受体,SRB为维持自身呼吸作用运转,能够产生导电性的“纳米导线”[25],能够以直接或间接的方式从铁基体获得电子,作为活性点提供电子而优先溶解,并且将红壤中Fe(III) 作为电子供体而还原[26,27]。尽管SRB胞外铁呼吸促发的铁氧化物还原的具体过程及机制尚不清晰,但该过程中SRB作为电子传输媒介,参与Fe和氧化铁间的电子转移,加速了Q235钢的腐蚀。
(1) 红壤中主要以赤铁矿和针铁矿形式富集的铁氧化物在腐蚀过程中作为阴极去极化剂,促进钢的腐蚀过程。
(2) 实验周期的前4 d为SRB的环境适应期,期间SRB对腐蚀电化学过程影响不显著;第4 d开始为SRB生长期,SRB呼吸代谢活动促使Q235钢的自腐蚀电位和极化电阻降低,腐蚀速率增大,在有菌红壤中的腐蚀速率约为在无菌红壤中的2倍。
(3) 细菌可与红壤颗粒表层铁氧化物发生作用,引起铁氧化物的微生物异化还原。其中,SRB作为电子传输媒介,参与Fe和FeOOH间的电子转移,这可能是促进Q235钢局部腐蚀电化学过程的主要原因。
The authors have declared that no competing interests exist.

/
| 〈 |
|
〉 |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}