


随着海洋经济的发展,作为探索和发展海洋经济的工具和平台,船舶及海洋工程日益增多。然而,海洋环境中微生物腐蚀严重危及它们的使用安全。微生物腐蚀作为海洋环境中腐蚀的一种重要的形式,指由微生物造成或影响的腐蚀,它威胁着人类生命安全和健康,并造成严重的经济损失[1,2]。数据指出,微生物腐蚀造成的损失约占总腐蚀损失的20%[3~6]。在与微生物腐蚀相关的众多的微生物中,硫酸盐还原菌(SRBs)是一种重要的类群[7~10]。这些微生物以硫酸盐为主要的电子受体,造成的腐蚀损失约占总微生物腐蚀损失的50%[11]。根据中国的腐蚀调查报告,中国由SRB腐蚀造成的经济损失达到了2000亿[12]。低合金是海洋工程领域中常用金属材料,由于SRB在海洋中的广泛分布,它极易受到SRB的腐蚀。
生物膜的形成依赖于趋化性为细菌提供导航的倾向[18,19],在鞭毛的运动下,它们会聚集到材料表面完成粘附[20]。然后细菌将在群体感应作用下完成生物膜的发育和成熟[21,22]。微生物膜的形成过程将直接受到外界环境的影响,而金属基质作为与细菌直接接触的部分,可以极大地影响微生物的附着和腐蚀。其中,合金元素会对细菌粘附产生重要影响,尤其是Cu,作为一种杀菌抑菌材料已被广泛研究[23~25]。然而,海洋工程材料,特别是合金钢,在设计时并没有考虑到微生物的影响,其中许多因素可能会影响微生物的附着和腐蚀,这一方面的研究很少。本文从常见的合金钢合金元素Mo入手,研究了Mo添加量对常见海洋腐蚀SRB的附着力和腐蚀性能的影响。阐述了Mo如何影响SRB粘附和腐蚀过程的分子机制,为今后厌氧微生物环境下海洋工程材料的设计和应用提供了科学依据。
1 实验方法
本实验采用合金钢作为钢样,由中国宝钢公司提供,其组成详见表1。对照钢(Cs)和含钼钢(Ms)样品采用线切割切割成10 mm × 10 mm × 3 mm的正方形样品块,然后用#50、#400、#800和#1200砂纸进行打磨。钢样用无水乙醇和丙酮超声清洗15 min,然后用氮气干燥。使用前在超净工作台上紫外灯下消毒1 h。
表1 对照钢(Cs)和含钼钢(Ms)的化学成分
Table 1
| Steel | C | Si | Mn | P | S | Nb | N | Ca | Mo | Fe |
|---|---|---|---|---|---|---|---|---|---|---|
| Cs | 0.05 | 0.2 | 1.5 | < 0.01 | < 0.001 | 0.02 | < 0.004 | 0.0025 | 0.0 | Bal. |
| Ms | 0.05 | 0.2 | 1.5 | < 0.01 | < 0.001 | 0.02 | < 0.004 | 0.0025 | 1.0 | Bal. |
本研究使用的SRB(Desulfovibrio vulgaris)购自American Type Culture Collection(ATCC),实验使用的培养基为修正的PGC培养基:0.5 g KH2PO4,1 g NH4Cl,0.06 g CaCl2·6H2O,0.06 g MgSO4·7H2O,6 mL70%乳酸钠,1 g酵母提取物,0.3 g柠檬酸钠放入1 L的陈海水中。刃天青作为培养液中的指示剂。所有试剂菌购自于国药集团化学试剂有限公司。接种前,将培养基在121 ℃高压灭菌20 min,冷却至室温((25 ± 2) ℃)后,以200 mL/min的速率用高纯度N2 (> 99.99%,体积分数)脱氧45 min,用0.22 μm滤膜将L-半胱氨酸溶液加入培养基中。最后用1 mol/L NaOH或1 mol/L HCl溶液调节培养基的pH至7.2~7.4。然后,将2 mL处于对数生长期的SRB菌种接种在厌氧手套箱(LAI-D2)中接种到含有400 mL培养基的500 mL广口瓶。然后用环氧树脂密封广口瓶,在37 ℃恒温摇床(SPM-508)中以120 r/min转速培养。
采用Gammy Interface 1010E电化学工作站来进行电化学阻抗谱(EIS)测试,用于电化学测试的三电极系统由样品作为工作电极,饱和甘汞电极作为参比电极,铂板电极作为对电极组成。测试溶液采用灭菌培养基,测试周期为14 d。样品在培养基中浸泡14 d后,按照ASTM G1-2003标准去除腐蚀产物。通过称重测试开始和结束时样品质量的差值来计算腐蚀速率,腐蚀速率计算公式为:Vcorr = (87600Δm)/ρAt,Vcorr、t、ρ、A和Δm分别代表腐蚀速率(mm/a),浸泡时间(h),材料密度(g/cm3),暴露面积(cm2)和质量损失(g)。
浸泡14 d后取出样品,用磷酸缓冲液(PBS,pH = 7.4)洗涤3次,然后在2.5%戊二醛溶液中浸泡2 h,固定表面微生物。然后用PBS洗涤3次,用不同浓度的乙醇(30%、50%、70%、80%、90%和100%)脱水,并用高纯度(99.999%)氮气干燥。采用扫描电子显微镜-能谱仪(SEM-EDS,JEOL JSM-7500 F)对腐蚀试样的表面和截面形貌进行了表征。腐蚀产物采用X射线衍射仪(XRD,PANalytica,2θ = 20~90°)表征。在倒置荧光显微镜(FM,Ti-S)下观察用吖啶橙荧光染色的样品上的微生物附着情况。将样品表面的腐蚀产物去除并在纯氮中干燥后,用光学轮廓仪(Bruker Contour GT)观察样品表面的点蚀,并进行统计分析。
合金钢表面形成生物膜,接种2 d后采集生物膜样品。从培养基中取出样品后,在0 ℃的0.85% (质量分数) NaCl缓冲液中快速轻洗,去除污染的悬浮细胞,然后在0 ℃的0.85%NaCl缓冲液中超声处理2 min,从金属表面收集生物膜细胞。将含有生物膜细胞的缓冲液在-2 ℃下离心3 min,沉淀的生物膜细胞在0 ℃下用6 mL 0.85%NaCl缓冲液重悬,转移到冷头搅拌管中,室温下离心15 s。然后,将细胞颗粒立即冷冻在干冰乙醇浴中,随后将样品送至上海美吉生物医药科技有限公司,进行转录组检测。
2 结果与讨论
2.1 合金钢表面细菌附着及生物膜形成分析
图1

图1
荧光显微镜下细菌在Cs和Ms表面附着情况
Fig.1
Fluorescence microscopes of adhesions of SRB on Cs (a) and Ms (b)
2.2 合金钢的表面形貌和成分分析
图2显示了合金钢在无菌培养基中浸泡14 d后表面的SEM图像。观察了到Ms表面的腐蚀形态与Cs相似,在厌氧环境下表面的腐蚀产物较少。
图2

图2
Cs和Ms在无菌培养基中浸泡14 d后表面SEM图像
Fig.2
SEM surface images of Cs (a) and Ms (b) after 14 d immersion in abiotic medium
图3

图3
Cs和Ms在有菌培养基中浸泡14 d后表面的SEM图像
Fig.3
SEM surface images of Cs (a) and Ms (b) after 14 d immersion in biotic medium
图4

图4
Cs和Ms 在有菌培养基中浸泡14 d后的SEM横截面图
Fig.4
SEM cross-sectional images of Cs (a) and Ms (b) after 14 d immersion in biotic medium
图5

图5
Cs和Ms在含D.vulgaris的培养基中浸泡14 d后的XRD谱图
Fig.5
XRD pattern of Cs (a) and Ms (b) after 14 d immersion in biotic medium containing D.vulgaris
图6

图6
Cs和Ms在有菌培养基中浸泡14 d后的EDS结果
Fig.6
EDS results of Cs (a) and Ms (b) after immersion in biotic medium for 14 d
2.3 合金钢表面点蚀分析
图7

图7
Cs和Ms在无菌培养基中浸泡14 d后点蚀的二维和三维图像
Fig.7
2D (a, b) and 3D (c, d) images of pitting of Cs (a, c) and Ms (b, d) after immersion in abiotic medium for 14 d
图8

图8
Cs和Ms在有菌培养基中浸泡14 d后点蚀的二维和三维图像
Fig.8
2D (a, b) and 3D (c, d) images of pitting of Cs (a, c) and Ms (b, d) after immersion in biotic medium for 14 d
图9显示了浸泡在无菌和有菌培养基中合金钢表面点蚀的统计图。可以看出,在无菌培养基中,Ms的表面点蚀直径和深度比Cs减小,这也证明了加入Mo后,腐蚀,特别是合金钢的点蚀略有减少。在D.vulgaris存在的情况下,Ms的腐蚀明显更严重,直径增加明显,但深度变化不显著。这也证明了添加Mo增加了合金钢在D.vulgaris存在下的腐蚀,使点蚀更加严重。此外,随着Mo的加入,点蚀的数量增加,这证实了Mo可以促进SRB的附着,从而加速合金钢的腐蚀。
图9

图9
Cs和Ms在无菌和有菌培养基中浸泡10 d后表面点蚀统计
Fig.9
Size statistics of corrosion pits of Cs和Ms after 14 d immersion in abiotic medium and biotic medium
2.4 失重和EIS测试分析
图10为合金钢在无菌和生物培养基中浸泡14 d后,去除表面腐蚀产物后计算的腐蚀速率。如图所示,在无菌培养基中,Mo对合金钢腐蚀速率的影响较小。在有菌培养基中,Ms和Cs的腐蚀情况有明显的区别,Ms的腐蚀速率明显高于Cs,这也证明了Mo的加入加速了合金钢在生物介质中的腐蚀速率。
图10

图10
Ms和Cs在无菌和有菌培养基中的腐蚀速率
Fig.10
Corrosion rates of Ms and Cs in abiotic and biotic mediums
图11

图11
Ms和Cs在无菌和有菌培养基中的Nyquist图和相应等效电路图
Fig.11
Nyquist plots of Ms (a, c) and Cs (b, d) in abiotic (a, b) and biotic (c, d) mediums, and equivalent circuit diagrams (e, f)
表2 Cs和Ms在无菌培养基中浸泡不同时间后的EIS的拟合参数
Table 2
| Group | Rs / Ω·cm2 | Yf / S·s n ·cm-2 | n | Rfilm / Ω·cm2 | Ydl / S·s n ·cm-2 | n | Rct / Ω·cm2 |
|---|---|---|---|---|---|---|---|
| Cs (1 d) | 60.79 | 6.8 × 10-5 | 0.9 | 3.6 × 102 | 2.3 × 10-4 | 0.7 | 5.2 × 103 |
| Cs (9 d) | 49.56 | 1.8 × 10-4 | 0.9 | 2.5 × 104 | 8.4 × 10-5 | 0.9 | 2.2 × 104 |
| Cs (14 d) | 56.02 | 8.0 × 10-5 | 1 | 2.4 × 104 | 7.7 × 10-5 | 0.9 | 6.3 × 104 |
| Ms (1 d) | 68.25 | 3.8 × 10-5 | 0.8 | 6.4 × 103 | 4.5 × 10-5 | 0.9 | 2.4 × 104 |
| Ms (9 d) | 69.96 | 10.0 × 10-5 | 0.9 | 5.9 × 103 | 1.4 × 10-3 | 0.5 | 2.9 × 102 |
| Ms (14 d) | 68.14 | 1.1 × 10-4 | 0.8 | 1.7 × 104 | 1.1 × 10-4 | 0.9 | 9.1 × 104 |
表3 Cs和Ms在有菌培养基中浸泡不同时间后的EIS拟合参数
Table 3
| Group | Rs / Ω·cm2 | Yf / S·s n ·cm-2 | n | Rfilm / Ω·cm2 | Ydl / S·s n ·cm-2 | n | Rct / Ω·cm2 |
|---|---|---|---|---|---|---|---|
| Cs (1 d) | 60.20 | 3.1 × 10-5 | 0.6 | 49.2 | 3.6 × 10-5 | 0.8 | 2.9 × 104 |
| Cs (9 d) | 65.70 | 1.3 × 10-3 | 0.4 | 9.7 × 102 | 3.1× × 10-4 | 0.9 | 1.5 × 103 |
| Cs (14 d) | 45.58 | 2.6 × 10-4 | 0.9 | 4.6 × 103 | 3.4 × 10-5 | 0.5 | 93.1 |
| Ms (1 d) | 64.50 | 1.7 × 10-4 | 0.9 | 23.2 | 2.9 × 10-4 | 0.9 | 8.2 × 103 |
| Ms (9 d) | 59.95 | 1.1 × 10-3 | 0.6 | 17.7 | 3.4 × 10-4 | 0.9 | 1.6 × 104 |
| Ms (14 d) | 61.08 | 3.1 × 10-4 | 0.7 | 22.3 | 2.0 × 10-4 | 0.9 | 6.0 × 104 |
2.5 RNA-seq分析
图12为Mo对合金钢表面D.vulgaris粘附腐蚀的分子调控机制。本文选择了一些差异显著的基因,如图12a所示。很明显,Mo的加入显著上调了参与细胞内S2-生成反应、鞭毛和趋化性的基因。在细胞内,细胞内腐蚀相关酶(dsrABC、aprAB、sat、por和ack)参与S2-生成反应[27]。如图12b所示,在D.vulgaris细胞中,许多酶参与了这一过程的共同调控,其中硫酸盐渗透酶(SulP)用于允许SO
图12
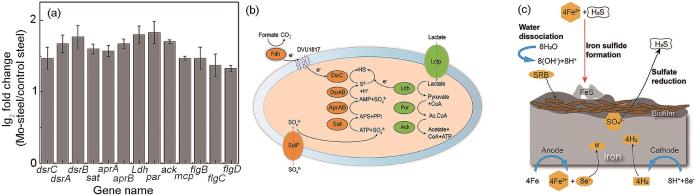
图12
Mo影响D.vulgaris在合金钢表面的附着和腐蚀的分子控制机理图
Fig.12
Schematic illustrations of molecular mechanism about the effect of Mo on the adhesion and corrosion of D.vulgaris on Ms: (a) gene diagram; (b) S2- synthesis related enzymes; (c) corrosion process
在阳极反应,Fe原子失去电子,变成Fe2+,
水电离成H+和OH-,
在阴极反应,H+得到电子,变成H原子
D.vulgaris介导SO
Fe2+和S2-在环境中生成FeS,和OH-生成Fe(OH)2
总反应如下:
阴极的去极化反应与D.vulgaris的去极化反应密不可分。D.vulgaris释放氢化酶,导致金属氧化H原子,阻止阴极的极化反应,导致阴极去极化。S与Fe2+的反应产生FeS,导致钢的阴极去极化,而阴极去极化引起H原子的不断损失,从而增加了Fe的腐蚀[7,28]。此外,一些控制鞭毛和趋化性的基因表达也发生了变化。flgBCD基因负责鞭毛基质杆,mcp基因控制细菌的趋化性。图12c为SRB的具体腐蚀过程,在试样表面形成膜后,介导H原子与硫酸盐反应生成H2S,H2S又反应生成硫铁氧体[29,30]。在这一过程中,Mo的存在影响了D.vulgaris细胞中不同途径的基因表达,从而加速了合金钢的腐蚀。
2.6 腐蚀机制
溶解在铁氧体中的Mo能产生固溶强化效应。Mo可以形成碳化钼,碳化钼在析出强化和晶粒细化中起着关键作用,可以提高材料的抗拉强度和屈服强度。在不锈钢中,Mo的加入也可以提高材料的耐腐蚀性。因此,Mo常被添加到合金材料中以改善其性能。然而,在海洋工程材料的设计中,并没有考虑合金元素对微生物腐蚀的影响。在本工作中,合金钢中加入Mo后,钢在无菌环境中的耐腐蚀性没有明显提高。相比之下,D.vulgaris的存在加速了合金钢的腐蚀。最后,通过RNA-seq,可见这正是由于Mo对细菌胞内S2-生成过程、趋化性和鞭毛基因的调控。其中,Mo增加了S2-的生成,提高了D.vulgaris在合金钢表面的附着力和成膜能力。在之前研究中,也观察到Mo可以影响解脂假交替单胞菌和铜绿假单胞菌的附着和随后的腐蚀行为[31,32]。在本研究中,鞭毛基因的上调也提高了D.vulgaris对材料表面的粘附和成膜能力,加速了其对合金钢表面的粘附[33~36]。在SRB中,鞭毛也可以进行电子传递,属于SRB的直接电子传递机制(DET)[37~40]。因此,除了生物膜的形成,Mo可能影响DET机制。与以往研究不同的是,对于D.vulgaris,Mo也会影响S2-的生产过程,在1.0% (质量分数) Mo时增加其产量,并且与生物膜形成的偶合因素共同加剧了材料的腐蚀。这可能是因为环境中钼的含量相对较低,但它也是生物体中许多酶和反应的必需元素。因此,当其含量高于环境含量时,细菌会在趋化性的引导下,利用鞭毛向带Mo的钢表面游去,形成更稳定、更致密的生物膜。
3 结论
(1) 研究表明合金钢中的Mo对SRB腐蚀有调节作用。在SRB存在的情况下,含Mo合金钢宏观腐蚀速率提升约40%,材料表面点蚀数目增多。
(2) 在Mo的作用下,SRB在含Mo钢表面附着量增加,表面含S腐蚀产物增多。相关转录组测试表明,参与SRB的S2-形成的部分基因表达增强,影响生物膜和DET的鞭毛基因和趋化基因也被上调,从而提高了细菌形成生物膜和还原SO
(3) 在进行材料设计时,应当考虑到使用环境中的微生物,避免因微生物腐蚀造成材料失效加速。
参考文献
Microbiologically influenced corrosion: an update
[J].
Laboratory investigation of microbiologically influenced corrosion of Q235 carbon steel by halophilic archaea Natronorubrum tibetense
[J].
Adaptive bidirectional extracellular electron transfer during accelerated microbiologically influenced corrosion of stainless steel
[J].
Review on bacterial biofilm: an universal cause of contamination
[J].
Microbiologically influenced corrosion (MIC) management using Bayesian inference
[J].
Study of corrosion behavior of copper in 3.5 wt.%NaCl solution containing extracellular polymeric substances of an aerotolerant sulphate-reducing bacteria
[J].
Synergistic effects of deposits and sulfate reducing bacteria on the corrosion of carbon steel
[J].
Influence of nutrition on Cu corrosion by Desulfovibrio vulgaris in anaerobic environment
[J].
SRB-mediated corrosion of marine submerged AISI 1020 steel under impressed current cathodic protection
[J].
Copper corrosion monitoring by electrical resistance probes in anoxic groundwater environment in the presence and absence of sulfate reducing bacteria
[J].
Mitigation of sulfate reducing Desulfovibrio ferrophilus microbiologically influenced corrosion of X80 using THPS biocide enhanced by Peptide A
[J].
The cost of corrosion in China
[J].
Mathematical modelling of microbial corrosion in carbon steel due to early-biofilm formation of sulfate-reducing bacteria via extracellular electron transfer
[J].
Inadequate dosing of THPS treatment increases microbially influenced corrosion of pipeline steel by inducing biofilm growth of Desulfovibrio hontreensis SY-21
[J].
Mechanistic modeling of biocorrosion caused by biofilms of sulfate reducing bacteria and acid producing bacteria
[J].
Biocorrosion is also known as microbiologically influenced corrosion (MIC). Most anaerobic MIC cases can be classified into two major types. Type I MIC involves non-oxygen oxidants such as sulfate and nitrate that require biocatalysis for their reduction in the cytoplasm of microbes such as sulfate reducing bacteria (SRB) and nitrate reducing bacteria (NRB). This means that the extracellular electrons from the oxidation of metal such as iron must be transported across cell walls into the cytoplasm. Type II MIC involves oxidants such as protons that are secreted by microbes such as acid producing bacteria (APB). The biofilms in this case supply the locally high concentrations of oxidants that are corrosive without biocatalysis. This work describes a mechanistic model that is based on the biocatalytic cathodic sulfate reduction (BCSR) theory. The model utilizes charge transfer and mass transfer concepts to describe the SRB biocorrosion process. The model also includes a mechanism to describe APB attack based on the local acidic pH at a pit bottom. A pitting prediction software package has been created based on the mechanisms. It predicts long-term pitting rates and worst-case scenarios after calibration using SRB short-term pit depth data. Various parameters can be investigated through computer simulation. Copyright © 2016 Elsevier B.V. All rights reserved.
Iron corrosion by novel anaerobic microorganisms
[J].
Inside Cover: biogenic iron sulfide nanoparticles to enable extracellular electron uptake in sulfate-reducing bacteria (Angew. Chem. Int. Ed. 15/2020)
[J].
Sensing and approaching toxic arsenate by Shewanella putrefaciens CN-32
[J].
Energy taxis toward redox-active surfaces decreases the transport of electroactive bacteria in saturated porous media
[J].
Coordinating assembly of a bacterial macromolecular machine
[J].The assembly of large and complex organelles, such as the bacterial flagellum, poses the formidable problem of coupling temporal gene expression to specific stages of the organelle-assembly process. The discovery that levels of the bacterial flagellar regulatory protein FlgM are controlled by its secretion from the cell in response to the completion of an intermediate flagellar structure (the hook-basal body) was only the first of several discoveries of unique mechanisms that coordinate flagellar gene expression with assembly. In this Review, we discuss this mechanism, together with others that also coordinate gene regulation and flagellar assembly in Gram-negative bacteria.
Staphylococcus quorum sensing in biofilm formation and infection
[J].
Reactor scale modeling of quorum sensing induced biofilm dispersal
[J].
Effect of surface passivation on corrosion resistance and antibacterial properties of Cu-bearing 316L stainless steel
[J].
Antibacterial ability of a novel Cu-bearing 2205 duplex stainless steel against Pseudomonas aeruginosa biofilm in artificial seawater
[J].
An investigation of the antibacterial ability and cytotoxicity of a novel cu-bearing 317L stainless steel
[J].In order to solve the challenging problem of microbial infections caused by microorganisms on medical implants, it is imperative to develop novel antimicrobial biomaterials. This work demonstrated that 317L-Cu stainless steel (SS), created by adding copper through a solution and aging heat treatment process, exhibited good antibacterial properties against staphylococcus aureus, achieving 2 log reduction of planktonic cells after 5 days of incubation. In this study, the antibacterial test was performed using the plate count method, the fluorescence cell staining method and the quantitative polymerase chain reaction (qPCR) method. It is well known that a high concentration of copper ion can lead to cytotoxicity. This work explored the cytotoxicity of 317L-Cu SS through real-time cell analysis (RTCA). Experimental results demonstrated that the 317L-Cu SS possessed a satisfactory antibacterial ability against S. aureus, and the antibacterial rate based on the reduction of sessile cell count reached 98.3% after 24-hour treatment. The bacterial adhesion and the biofilm thickness were considerably reduced by the 317L-Cu SS. The results of RTCA suggested that 317L-Cu SS did not introduce cytotoxicity to mouse cells, indicating its suitability as a medical implant material.
Synergistic inhibition of rhamnolipid and 2,2-dibromo-3-hypoazopropionamide on microbiologically influenced corrosion of X80 pipeline steel
[J].
鼠李糖脂与2,2-二溴-3-次氮基丙酰胺协同抑制X80管线钢的微生物腐蚀
[J].研究了2,2-二溴-3-次氮基丙酰胺(DBNPA)与鼠李糖脂(RL)对X80管线钢在硫酸盐还原菌Desulfovibrio bizertensis SY-1中的腐蚀行为影响。结果表明,Desulfovibrio bizertensis SY-1存在时,X80管线钢的腐蚀失重和点蚀深度明显增加,且表面检测出FeS腐蚀产物。DBNPA的添加,抑制了浮游及固着SRB的生长,减缓了X80管线钢的腐蚀。150 mg/L DBNPA与500 mg/L RL进行复配时,X80管线钢的腐蚀速率与SRB体系相比降低了77.8% (p = 0.009),与单独使用300 mg/L DBNPA相比降低了50%。另外,150 mg/L DBNPA与500 mg/L RL复配时,X80管线钢腐蚀浸泡15 d后的腐蚀电流密度与SRB体系相比降低了84.7%,与单独使用300 mg/L DBNPA的SRB体系相比降低了20.5%,可显著抑制X80管线钢的微生物腐蚀。
Gene sets and mechanisms of sulfate-reducing bacteria biofilm formation and quorum sensing with impact on corrosion
[J].
Corrosion of carbon steel by sulphate reducing bacteria: initial attachment and the role of ferrous ions
[J].
Spectroscopic study on biological mackinawite (FeS) synthesized by ferric reducing bacteria (FRB) and sulfate reducing bacteria (SRB): Implications for in-situ remediation of acid mine drainage
[J].
Rapid pyritization in the presence of a sulfur/sulfate-reducing bacterial consortium
[J].Sedimentary pyrite (FeS) is commonly thought to be a product of microbial sulfate reduction and hence may preserve biosignatures. However, proof that microorganisms are involved in pyrite formation is still lacking as only metastable iron sulfides are usually obtained in laboratory cultures. Here we show the rapid formation of large pyrite spherules through the sulfidation of Fe(III)-phosphate (FP) in the presence of a consortium of sulfur- and sulfate-reducing bacteria (SRB), Desulfovibrio and Sulfurospirillum, enriched from ferruginous and phosphate-rich Lake Pavin water. In biomineralization experiments inoculated with this consortium, pyrite formation occurred within only 3 weeks, likely enhanced by the local enrichment of polysulfides around SRB cells. During this same time frame, abiotic reaction of FP with sulfide led to the formation of vivianite (Fe(PO)·8HO) and mackinawite (FeS) only. Our results suggest that rates of pyritization vs. vivianite formation are regulated by SRB activity at the cellular scale, which enhances phosphate release into the aqueous phase by increased efficiency of iron sulfide precipitation, and thus that these microorganisms strongly influence biological productivity and Fe, S and P cycles in the environment.
Molybdenum-mediated chemotaxis of Pseudoalteromonas lipolytica enhances biofilm-induced mineralization on low alloy steel surface
[J].
Pseudomonas aeruginosa-accelerated corrosion of Mo-bearing low-alloy steel through molybdenum-mediating chemotaxis and motility
[J].
Geobacter metallireducens accesses insoluble Fe(III) oxide by chemotaxis
[J].
Bacterial flagella explore microscale hummocks and hollows to increase adhesion
[J].Biofilms, surface-bound communities of microbes, are economically and medically important due to their pathogenic and obstructive properties. Among the numerous strategies to prevent bacterial adhesion and subsequent biofilm formation, surface topography was recently proposed as a highly nonspecific method that does not rely on small-molecule antibacterial compounds, which promote resistance. Here, we provide a detailed investigation of how the introduction of submicrometer crevices to a surface affects attachment of Escherichia coli. These crevices reduce substrate surface area available to the cell body but increase overall surface area. We have found that, during the first 2 h, adhesion to topographic surfaces is significantly reduced compared with flat controls, but this behavior abruptly reverses to significantly increased adhesion at longer exposures. We show that this reversal coincides with bacterially induced wetting transitions and that flagellar filaments aid in adhesion to these wetted topographic surfaces. We demonstrate that flagella are able to reach into crevices, access additional surface area, and produce a dense, fibrous network. Mutants lacking flagella show comparatively reduced adhesion. By varying substrate crevice sizes, we determine the conditions under which having flagella is most advantageous for adhesion. These findings strongly indicate that, in addition to their role in swimming motility, flagella are involved in attachment and can furthermore act as structural elements, enabling bacteria to overcome unfavorable surface topographies. This work contributes insights for the future design of antifouling surfaces and for improved understanding of bacterial behavior in native, structured environments.
Integrated control of surface adaptation by the bacterial flagellum
[J].Many bacteria can alternate between motile and sessile lifestyles, and wide-ranging sets of environmental stimuli regulate the transition from a free-swimming to a surface-attached state. A transenvelope machine called the flagellum, known primarily for its role in promoting cellular motility, stimulates the motile-sessile transition by detecting contact with solid substrates. Recent work has revealed a striking level of sophistication within the regulatory circuits that link flagellar function to surface colonization. I describe the current paradigm whereby the flagellum promotes the sessile state by increasing production of the second-messenger bis-(3'-5')-cyclic diguanosine monophosphate (c-di-GMP). I then highlight studies that have identified multiple routes by which the flagellum activates c-di-GMP production, calling the concept of a linear surface recognition pathway into the question. I conclude by proposing a role for the flagellum as a signaling hub that integrates environmental stimuli to coordinate a surface colonization program that occurs across a range of spatial and temporal scales.Copyright © 2021. Published by Elsevier Ltd.
An inside look at a biofilm: Pseudomonas aeruginosa flagella biotracking
[J].
Flagella act as Geobacter biofilm scaffolds to stabilize biofilm and facilitate extracellular electron transfer
[J].
Flagellum mediates symbiosis
[J].We report here molecular mechanisms underlying a bacteria-archaeon symbiosis. We found that a fermentative bacterium used its flagellum for interaction with a specific methanogenic archaeon. The archaeon perceived a bacterial flagellum protein and activated its metabolism (methanogenesis). Transcriptome analyses showed that a substantial number of genes in the archaeon, including those involved in the methanogenesis pathway, were up-regulated after the contact with the flagellum protein. These findings suggest that the bacterium communicates with the archaeon by using its flagellum.
Electrically conductive bacterial nanowires produced by Shewanella oneidensis strain MR-1 and other microorganisms
[J].
Deteriorated biofilm-forming capacity and electroactivity of Shewanella oneidnsis MR-1 induced by insertion sequence (IS) elements
[J].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}