


海洋工程装备和船舶用钢的微生物腐蚀 (MIC) 问题每年给国家造成约千亿元的经济损失,亟待关注和解决[1-3]。微生物腐蚀按照微生物所处的环境可分为厌氧微生物腐蚀和好氧微生物腐蚀。以往的研究更多关注厌氧微生物腐蚀,比如硫酸盐还原菌 (SRB) 所造成的腐蚀,其腐蚀机理主要是阴极去极化理论和电子传递理论。但海洋环境中好氧微生物腐蚀所涉及的腐蚀性细菌种属较多,其腐蚀机理的探讨主要围绕生物膜造成的氧浓差腐蚀和胞外分泌物 (EPS) 与金属离子的络合作用[4-6]。随着微生物学、海洋化学和材料学等相关学科领域的日益交叉和融合,研究[7]表明,海洋细菌分泌的活性次级代谢产物 (ASM) 如抗生素、激素、色素等也可能会影响金属腐蚀过程,其中细菌色素对细菌的生长发育没有显著的作用,却具有一定的理化性质和生物学功能[8]。很多海洋产色菌分泌的色素物质是具有杂化芳香结构的有机分子,含有氧化还原性质的功能基团,可以作为内源性的氧化还原介体在细菌和材料之间传递电子并发生氧化还原反应,这类色素物质包括小分子的有机化合物,如吩嗪、绿脓素、醌类、灵菌红素等[9];也有少数是大分子有机化合物,如黑色素[10]。而金属腐蚀的本质就是金属在电解质溶液中电子的得失与传递,因此海洋产色菌与色素物质将很可能参与到金属材料的腐蚀过程中并影响其腐蚀行为。假交替单胞菌属 (Pseudoalteromonas) 是一种典型的海洋易产色菌属,在种属内分为产色 (Pigmented) 和不产色 (Non-Pigmented) 菌种,且易于在各种材料表面附着形成生物膜[11]。
本文中P. lipolytica的产色基因突变菌株ΔhmgA能够在生长周期内分泌产生黑色素,这是一种广泛存在于生物体内的高分子醌类聚合物,由于其独特的非晶半导体性质,既可以作为细菌胞外呼吸的电子受体,又可以作为氧化还原介体行使电子传递载体的功能[12]。316L不锈钢由于其优异的自钝化性能和耐腐蚀性能成为海洋工程关键部件的首选材料之一。316L不锈钢的自钝化行为是由于不锈钢中的Cr较高,当环境中含有氧时,会形成一层几十个纳米厚的钝化膜,这种钝化膜不但具有很好的防腐性能,且可以进行自我修复,从而保护金属不被腐蚀。但当这层钝化膜被破坏后而又无法自修复时,不锈钢也会发生点蚀和缝隙腐蚀[13-16]。以往的研究大多认为海水中的Cl-是不锈钢发生点蚀的“罪魁祸首”。当不锈钢所在的服役环境中含有能够分泌色素物质的P. lipolytica时,黑色素物质是否会破坏不锈钢的钝化膜,而不锈钢是否依然可以保持良好的耐蚀性能,需要进一步研究和探索。
针对以上的问题,本实验选用了361L不锈钢作为腐蚀试验材料,以P. lipolytica的野生菌株 (WT) 与产色菌株 (∆hmgA) 为模式菌株,重点研究海洋细菌分泌黑色素加速不锈钢钢腐蚀的行为和机理,研究在含有产色细菌和黑色素物质的环境条件下不锈钢的腐蚀机理以及黑色素对钝化膜形成和点蚀发展的影响机制,从而丰富和拓展微生物腐蚀机理。
1 实验方法
1.1 样品制备
本文所用不锈钢样品为宝钢中央研究院提供的海洋工程用316L不锈钢,其成分 (质量分数,%) 为:Mn 1.24,Ni 10.12,Mo 2.05,Cr 16.54,C 0.03,Si 0.52,S 0.003,P 0.03,余量Fe。不锈钢试样采用线切割加工成尺寸为10 mm×10 mm×3 mm的块体,其中电化学测试样品背面焊接金属导线,用环氧树脂进行封装使其仅露出10 mm×10 mm工作面。样品采用SiC砂纸逐级打磨至1200#,后放入无水乙醇溶液内超声清洗5 min,再分别用丙醇和去离子水清洗除油后干燥备用。实验前所有不锈钢样品需紫外照射至少0.5 h以进行灭菌。
1.2 细菌培养及生理生化表征
本文使用的菌种为中科院南海所王晓雪研究员提供的海洋解脂假交替单胞菌P. lipolytica SCSIO 04301,野生菌株 (WT) 分离自南海,分离海域水温约为24~29 ℃,盐度约为33.2‰~34.2‰,并完成了全基因序列测试[17]。WT在基因簇hmgA发生点突变后形成了能够分泌黑色素的基因突变株∆hmgA,与野生菌株共同构成本文中研究海洋微生物腐蚀的模式菌。
本文中使用来自青岛海博的2216E模拟海水液体培养基对细菌进行培养,使用高压灭菌锅对配置完成后的培养基高温高压灭菌得到无菌2216E海水培养基。实验中用到的培养基均为灭菌后无菌培养基。分别将P. lipolytica的WT和∆hmgA接种至100 mL培养基中培养得到细菌培养液。用无菌培养基调节,使得细菌培养基中细菌浓度约为106 CFU/mL。取100 μL调节后细菌溶液至200 mL培养基中,置于恒温 (37 ℃) 摇床中振荡 (120 r/min) 培养,获得∆hmgA细菌发酵液。另取菌液于试管中,25 ℃静置培养3 d后在气液交界处获得生物被膜,收集后用灭菌去离子水稀释10倍后涂平板,再放置于25 ℃培养箱中直至长出细菌菌落进行形态学观察并拍照。
将WT和∆hmgA细菌发酵液设置3个平行实验组,通过标准平板活菌计数法每间隔1 d检测两种细菌的群落总数CFU/mL并绘制生长曲线[18]。同时在7 d培养过程中,每间隔1 d使用pH值计测量其pH值,使用溶解氧测定仪测量其溶解氧 (DO),并对每个实验组测得的数据进行平均数与方差计算,以绘制不同时间各细菌培养液的pH和溶解氧 (DO) 变化曲线。
1.3 细菌黑色素溶液的制备
将∆hmgA接种至灭菌培养基,同时添加1.5 g/L的酪氨酸底物以增加细菌黑色素分泌的浓度。将∆hmgA培养液置于培养箱内恒温 (37 ℃) 振荡培养 (120 r/min) 10 d获得黑色细菌发酵液。取细菌发酵液于低温高速离心机中,设置离心机温度为4 ℃,转速为10000 r/min,离心20 min,重复2~3次。取其上清液用规格为0.22 μm针筒过滤去除菌体,最后获得黑色素溶液,密封冷藏待用。
1.4 316 L不锈钢表面腐蚀形貌分析
分别将316L不锈钢试样浸泡于灭菌2216E海水培养基、WT培养液、∆hmgA培养液和黑色素溶液中。在37 ℃,转速为120 r/min恒温培养箱中振荡培养。14 d后,取出样品,用pH值为7.4的磷酸盐 (PBS) 缓冲溶液轻轻冲洗去除表面悬浮物,用LIVE/DEAD染色试剂盒对不锈钢样品进行染色标记,通过TCS SP8 STED 3X激光扫描共聚焦显微镜 (LSCM) 观察样品生物膜细菌活性状态;取出浸泡一定时间的不锈钢样品,用PBS缓冲溶液冲洗3次,后置于2.5%的戊二醛溶液固定细菌约2 h,后在一系列乙醇溶液 (30%、40%、50%、60%、70%、80%、90%、100%,乙醇体积分数) 浸泡15 min进行脱水处理,干燥后在扫描电镜 (SEM,JEOL JSM-7500F) 下观察其表面腐蚀形貌;使用X'Pert PRO XRD X射线衍射仪 (XRD) 对浸泡14 d后样品的腐蚀产物进行物相分析,PBS缓冲液轻轻冲洗浸泡后的样品表面,干燥后进行测试分析;通过Contour GT-X3白光干涉仪观察样品表面点蚀坑形貌及深度。观测前,根据NACE 0799方案修改版 (NACE,1999) 对样品表面进行预处理。取出浸泡14 d的不锈钢样品,依次浸泡于N,N-二丁基硫脲与盐酸混合溶液、饱和碳酸氢钠溶液、去离子水和丙酮溶液中2 min去除表面微生物及腐蚀产物,干燥后测试观察。
1.5 电化学分析测试
电化学测试采用三电极系统,其中不锈钢作为工作电极、铂片作为对电极、饱和甘汞电极作为参比电极。所有测试均由Metrohm电化学工作站完成,测试项目包括电化学阻抗 (EIS) 及线性极化电阻 (LPR) 测试。每次测试前,将测试体系在开路状态下静置30 min,直至开路电位趋于稳定。实验周期为14 d,电化学阻抗测试时间分别为第1、3、7、10、14 d,频率范围为105~10-2 Hz,正弦扰动幅值为±5 mV。线性极化电阻测试在第14 d测完电化学阻抗后测量,扫描速率为0.167 mV/s,扫描区间为-10~10 mV vs. OCP。实验数据经由电化学分析软件ZSimpWin 3.10拟合得到各项电化学参数。
2 结果与讨论
从图1a可以看出,WT和∆hmgA菌株在7 d内,生长总体保持一致 (CFU均保持在106数量级)。但是WT菌株在生长初期增殖比∆hmgA菌株快,在2 d达到指数生长期峰值,随后逐渐下降,∆hmgA则在3 d达到了指数生长期的峰值,而后逐步降至较低水平。从生长周期内的总体情况来看,能够分泌黑色素的基因突变株∆hmgA的生长活性没有出现比野生菌WT减弱的情况,这对于后续腐蚀性能的研究非常重要,说明两种细菌在活性上并没有显著差别。从图1b可以看出,WT和∆hmgA菌株在7 d内的pH值都呈现上升趋势。其中WT菌株培养液pH值从7.4升高至8.2,其中2 d到4 d升高比较平缓,从4 d开始升高较快;∆hmgA菌株培养液pH值从7.5升高至8.2。两种细菌溶液pH值的变化趋势基本一致。从1(C) 溶解氧的变化结果可以看出,7 d内WT和∆hmgA菌株培养液的溶解氧 (DO) 非常小,基本都在0.2 mol/L以下波动。这主要是由于两种菌株都是需氧型的,其新陈代谢会消耗溶液中的氧气,也就是说溶液中的氧含量对腐蚀的影响较小。
图1
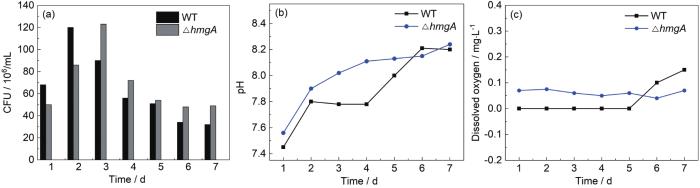
图1
P. lipolytica的野生菌 (WT)、基因突变株 (∆hmgA) 生长曲线,pH值,溶解氧在7 d内随时间变化规律
Fig.1
Changed with time of marine broths containing WT strain and mutant strain ∆hmgA in the culture cycle: (a) growth curve, (b) pH, (c) concentration of dissolved oxygen value
图2为316L不锈钢在不同溶液中浸泡14 d后的宏观照片。图2a所示不锈钢在无菌培养基溶液中浸泡14 d后表面仍然呈现出金属光泽,没有出现肉眼可见的腐蚀孔和腐蚀产物。图2b所示,不锈钢在WT培养液中浸泡14 d后,表面的金属光泽也非常明显,没有出现明显的点蚀和腐蚀产物。但在∆hmgA培养液中的不锈钢样品经过14 d浸泡后,表面则完全失去了金属光泽,并呈现均匀的灰白色 (图2c);在黑色素溶液中浸泡14 d后的不锈钢样品表面失去金属光泽,且出现较多锈蚀点。这两组中的不锈钢形貌与前两组相比,差异较为明显,这可能是因为P. lipolytica的产色菌及其黑色素对金属具有腐蚀作用,但具体腐蚀原因还需要进一步微观表征。
图2

图2
不锈钢样品在不同溶液中浸泡14 d后的宏观形貌
Fig.2
Macroscopic morphology of stainless-steel samples after immersion for 14 d: (a) abiotic, (b) WT strain, (c) ΔhmgA, (d) pyomelanin solution
为了表征WT和∆hmgA在不锈钢表面形成的生物膜,分别将浸泡1、7和14 d的不锈钢试样取出染色后通过LSCM进行表征。从图3可以看出,WT和∆hmgA菌株14 d内在不锈钢表面形成的生物膜中大部分是活菌细胞,这是因为不锈钢表面没有形成明显的腐蚀产物层,因此可以给予生物膜内细菌生存所必须的氧气和营养物质。从生物膜的形貌来看,WT与∆hmgA没有明显区别,但∆hmgA所形成的生物膜有先增厚后减薄的趋势,第7 d的生物膜厚度达到约62 µm,随后减薄到约42 µm。而WT所形成的生物膜的厚度一直是缓慢增加,第7 d后增加非常缓慢,从32 µm增厚到35 µm。生物膜厚度的区别可能与细菌诱导产生的沉积物有关,我们接下来将通过SEM与XRD进行进一步分析。
图3
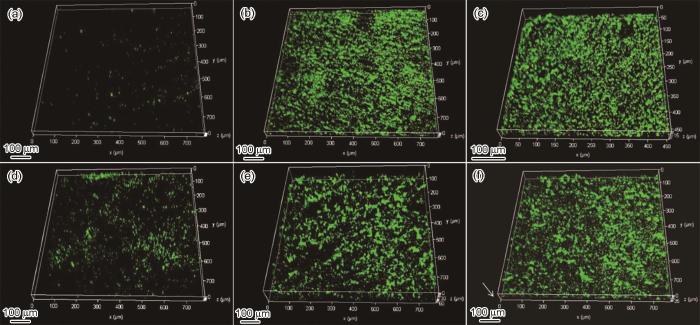
图3
不锈钢样品分别在WT和∆hmgA培养液中浸泡不同时间并染色标记后的CLSM图像
Fig.3
CLSM images of stainless steel samples immersed in WT (a-c) and ∆hmgA (d-f) medium for 1 d (a, d), 7 d (b, e) and 14 d (c, f), red and green represent dead and live cells, respectively
图4为不锈钢样品在不同细菌实验组中浸泡14 d后的SEM形貌。图4a为无菌对照组不锈钢的SEM图,其表面未见明显腐蚀产物,从放大图4b可以看到一些划痕,这是试样在打磨过程中造成的。图4c为WT溶液中不锈钢样品的SEM图,可以看出其表面没有明显的裂缝或腐蚀坑,但不均匀地黏附了一些胞状凸起物质,从放大图4d可以看到凸起上附着有很多细菌,推测为培养基中的有机物的黏附。图4e为∆hmgA细菌培养液中不锈钢样品的SEM图,可以明显看出样品表面覆盖一层致密的膜,且覆盖一些大小不均等的结晶颗粒物,放大图4f显示该膜层为紧密排布的矿物颗粒,矿物颗粒呈现三角锥体,且彼此之间存在一定缝隙,为典型的生物矿化膜表面形态。还有一些三角锥体状矿物颗粒团聚在一起形成花朵状矿物结晶。课题组前期研究工作中曾发现P. lipolytica由于过量分泌纤维素多糖,能够在低合金钢表面形成一层致密的生物矿化膜,该矿化膜是一种非常致密均匀的碳酸钙镁盐,从晶体形状和红外、XRD等检测结果判断,属于方解石结构[19]。∆hmgA只在基因簇hmgA发生了点突变,其他生理生化性质与WT是一致的,也能够正常分泌胞外多糖。但是不锈钢样品在WT菌液中没有发生矿化,而在∆hmgA菌液中出现了典型的生物矿化膜层,还需要进一步的实验研究。图4g和h为黑色素溶液中不锈钢钢样品的SEM图,可以看出样品表面的腐蚀产物不明显,表面也没有形成矿化产物,但由于黑色素溶液是经过细菌菌液过滤而来的,因此表面会附着一些死亡的细菌细胞。
图4

图4
不锈钢样品分别在不同溶液浸泡14 d后的表面生物膜形貌
Fig.4
Surface biofilm morphology of the 316L stainless steel surface in the abiotic (a, b), WT strain (c, d), ∆hmgA (e, f), pyomelanin media (g, h) after 14 d of immersion
如图5所示,通过XRD分析不锈钢样品在4种溶液中浸泡14 d后的表面产物成分。结果显示4组中的不锈钢样品都有2个主要不锈钢的峰,分别为43.5°和50.7°。另外,在∆hmgA菌液中还出现了29.8°,47.7°和48.7°的3个峰,这是碳酸钙沉淀的主要特征峰,表明∆hmgA能够在不锈钢样品表面产生了碳酸钙沉淀,这一结果与扫描电镜SEM图像是对应的。
图5

图5
不锈钢样品不同溶液内浸泡14 d后XRD谱
Fig.5
XRD spectra of stainless-steel samples immersed in different solutions for 14 d
图6为不同组浸泡14 d后不锈钢样品的原位白光干涉图像。其中图6a为无菌空白对照组的白光干涉图像,样品表面未见明显的点蚀;图6b为WT菌液中不锈钢样品的白光干涉图像,表面在划痕处出现了个别点蚀,但总体情况较好。说明在无菌和WT菌液中不锈钢由于钝化膜的存在有效抑制了点蚀。图6c可见,不锈钢在∆hmgA溶液中浸泡14 d后表面出现大量的点蚀坑,通过仪器所带的统计软件,可知点蚀深度约为17 μm。图6d为不锈钢在黑色素溶液中的白光干涉图像,表面也出现少量点蚀,以上结果显示产色菌及其黑色素可能是造成不锈钢点蚀的主要因素,黑色素是一种高分子醌类聚合物,它的醌基团具有可逆的氧化还原能力,这种能力能够破坏不锈钢的钝化膜,使得不锈钢处于活化状态,加速得失电子,从而引发腐蚀行为[19]。另外,不锈钢样品虽然能够在∆hmgA溶液中生成一层碳酸钙矿化膜,但这层矿化膜没有起到腐蚀防护的作用,相反,与黑色素溶液相比,矿化膜的形成还加剧了不锈钢的腐蚀程度,其机理需要通过电化学进一步探索和研究。
图6
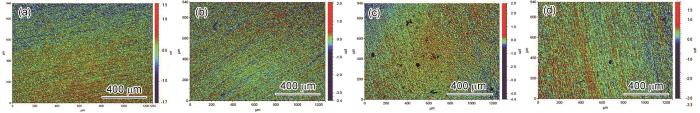
图6
不锈钢样品分别在无菌溶液、WT培养液、∆hmgA培养液和黑色素溶液浸泡14 d后白光干涉图像
Fig.6
Optical profilometry images of the pit morphologies of the 316 L stainless steel in Abiotic (a), WT strain (b), ∆hmgA (c), pyomelanin solution (d) after 14 d of immersion
利用动电位极化曲线可对不锈钢样品在四种溶液中的电化学动力学行为和钝化行为进行表征。从图7可以看出,除了在∆hmgA培养液中,其他3种溶液中不锈钢样品都出现了明显的钝化区,无菌溶液和WT培养液中不锈钢的维钝电位 (Ep) 约为-250 mV左右,钝化区间约为750 mV;在黑色素溶液中不锈钢的维钝电位正移约150 mV,为-100 mV,从维钝电位正移可知,黑色素的添加可能会延迟钝化现象的发生,使得不锈钢的钝化更加困难。但当不锈钢试样浸泡于∆hmgA菌液中时,阳极钝化区几乎消失,说明在此溶液中不锈钢无法发生钝化,结合扫描电镜照片和XRD图谱可知,不锈钢表面形成了矿化膜,矿化膜的存在使得钝化膜无法生成,阳极钝化区消失,阳极腐蚀电流密度急剧上升,导致腐蚀加速。从以上结果可以看出,当不锈钢浸泡到∆hmgA培养液中时,由于矿化产物和细菌色素的双重作用,含氧吸附粒子无法在不锈钢表面均匀吸附,从而导致钝化膜难以形成,而黑色素的存在使得点蚀非常严重。
图7
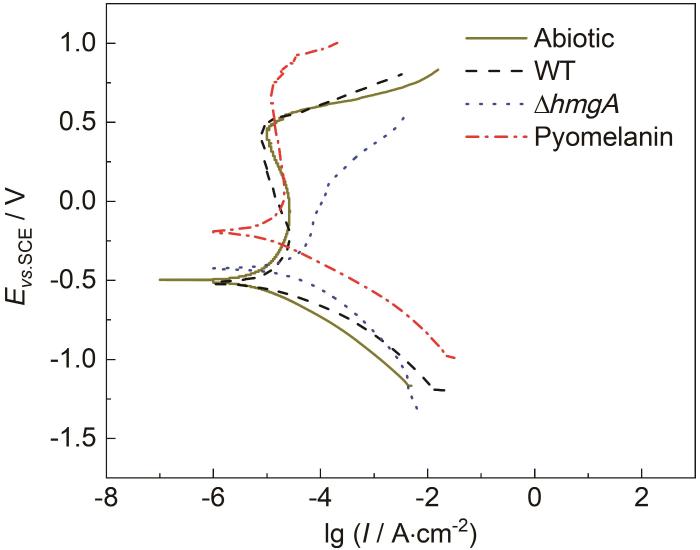
图7
不锈钢样品在不同溶液浸泡14 d后的动电位极化曲线
Fig.7
Potentiodynamic polarization curves of stainless-steel samples immersed in different solutions for 14 d
表1 不锈钢样品在不同溶液内浸泡不同时间后极化曲线的拟合参数
Table 1
| Solution time / d | Ecorr / V | Icorr / A·cm-2 | Ep / V |
|---|---|---|---|
| Abiotic-14 | -0.49 | 4.60×10-7 | -0.20 |
| ∆hmgA-14 | -0.42 | 1.13×10-6 | --- |
| WT-14 | -0.51 | 7.16×10-7 | -0.25 |
| Pyomelanin-14 | -0.19 | 8.46×10-7 | -0.30 |
通过电化学阻抗谱的Nyquist图也可以看出不锈钢在不同溶液中14 d内的电化学阻抗的变化趋势。从图8可以看出,除了∆hmgA以外,其他三种溶液中不锈钢的阻抗弧半径都非常大,并且呈现一个开放的趋势,是典型的不锈钢的阻抗特征,都可以用一个时间常数拟合,等效电路为Rs(RctQ)。在无菌溶液中,14 d内不锈钢的电荷转移电阻Rct的阻抗值由3×106 Ω·cm2逐渐升高到4.8×107 Ω·cm2,说明钝化膜在无菌海水中一直具有优良的保护作用。同样在WT培养液中,电化学阻抗值Rct在前3 d约为3×105 Ω·cm2,从第3 d开始,阻抗值逐渐增加,在14 d时增加到约3.7×107 Ω·cm2。而在黑色素溶液中,阻抗值Rct先增大后减小,第1 d阻抗值为5.4×106 Ω·cm2,第10 d逐渐升高到1.8×108 Ω·cm2,第14 d则降到3.5×106 Ω·cm2。在∆hmgA培养液中,钝化膜的阻抗谱特征消失,从第7 d开始出现了两个时间常数,拟合电路变为Rs(RctQ1)(RfQ2)。其中,膜电阻Rf的出现代表了不锈钢表面矿化产物膜的形成。从拟合结果来看,不锈钢在∆hmgA培养液中浸泡1 d后的Rct仅有1.8×103 Ω·cm2,即便表面生成矿化产物膜后也仅仅增加到3.5×104 Ω·cm2。从以上结果可以看出,由于产色菌在不锈钢表面可以形成矿化膜,因此使得不锈钢的电化学阻抗下降了至少2个数量级,尤其在前2 d,电化学阻抗下降了3个数量级,说明在没有形成矿化之前,钝化膜就已经被破坏了。细菌在材料表面形成完整的生物矿化膜,首先要在表面形成很多矿化位点,这些矿化位点的形成就开始破坏钝化膜,而不需要等到矿化膜完全形成。另外,不锈钢在浸泡初期,由于细菌色素的分泌使得钝化膜无法均匀覆盖在材料表面,因此为矿化成核提供了足够的位点,随着浸泡时间的延长,矿化膜才能在不锈钢表面形成,这同时也说明了为什么WT无法在不锈钢表面形成矿化膜,因为WT溶液中没有细菌黑色素,一旦钝化膜形成,生物矿化将失去足够的成核位点,因此无法在不锈钢表面形成生物矿化膜。
图8
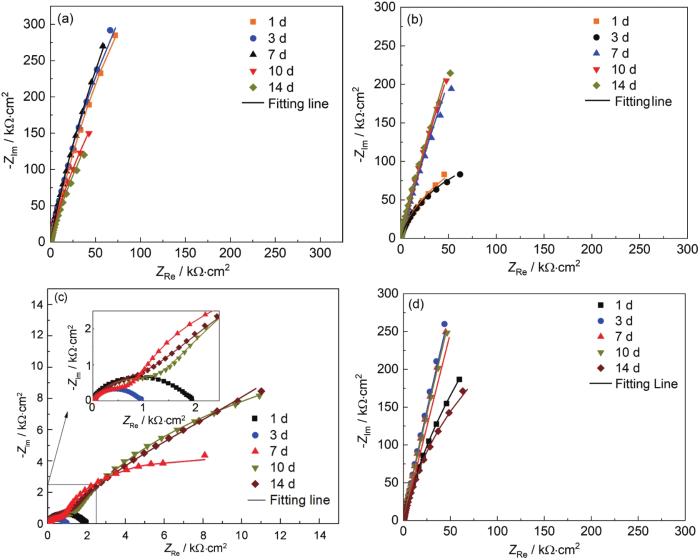
图8
不锈钢样品在不同溶液内浸泡不同时间后电化学阻抗谱
Fig.8
Electrochemical impedance spectroscopy of the 316 L stainless steel immersed in Abiotic (a), WT strain (b), ∆hmgA (c), pyomelanin solution (d) for different times
表2 不锈钢样品在不同溶液内浸泡不同时间后电化学阻抗拟合参数
Table 2
| Solution | Time / d | Rs / Ω·cm2 | Y1 / S·sec n ·cm-2 | n | Rf / Ω·cm2 | Y2 / S·sec n ·cm-2 | n | Rct / Ω·cm2 |
|---|---|---|---|---|---|---|---|---|
| Abiotic | 1 | 2.9 | --- | --- | --- | 4.0×10-5 | 0.9 | 3.0×106 |
| 3 | 4.5 | --- | --- | --- | 4.0×10-5 | 0.9 | 4.8×106 | |
| 7 | 6.4 | --- | --- | --- | 4.3×10-5 | 0.9 | 6.9×106 | |
| 10 | 3.4 | --- | --- | --- | 7.5×10-5 | 0.9 | 3.0×106 | |
| 14 | 5.8 | --- | --- | --- | 8.9×10-5 | 0.8 | 4.8×107 | |
| WT | 1 | 4.9 | --- | --- | --- | 3.4×10-5 | 0.8 | 3.1×105 |
| 3 | 5.3 | --- | --- | --- | 2.6×10-5 | 0.9 | 2.7×105 | |
| 7 | 6.6 | --- | --- | --- | 1.6×10-5 | 0.9 | 3.2×106 | |
| 10 | 9.5 | --- | --- | --- | 1.2×10-5 | 0.9 | 5.2×106 | |
| 14 | 8.5 | --- | --- | --- | 4.7×10-5 | 0.9 | 3.7×107 | |
| ∆hmgA | 1 | 3.6 | --- | --- | --- | 3.3×10-4 | 0.8 | 1.8×103 |
| 3 | 4.5 | --- | --- | --- | 6.4×10-4 | 0.8 | 9.8×102 | |
| 7 | 3.7 | 5.2×10-5 | 0.8 | 818 | 4.0×10-4 | 0.8 | 1.3×104 | |
| 10 | 1.8 | 3.4×10-5 | 0.6 | 872 | 3.1×10-4 | 0.9 | 3.5×104 | |
| 14 | 1.9 | 2.9×10-5 | 0.7 | 714.7 | 4.7×10-4 | 0.9 | 2.8×104 | |
| Pyomelanin | 1 | 8.2 | --- | --- | --- | 5.7×10-5 | 0.9 | 5.4×106 |
| 3 | 2.9 | --- | --- | --- | 6.4×10-4 | 0.8 | 2.7×107 | |
| 7 | 9.5 | --- | --- | --- | 8.7×10-4 | 0.6 | 1.7×107 | |
| 10 | 8.5 | --- | --- | --- | 4.7×10-4 | 0.9 | 1.8×107 | |
| 14 | 8.2 | --- | --- | --- | 6.0×10-5 | 0.8 | 3.5×106 |
从以上结果可以看出,如果钝化膜保持完好,没有被破坏,即使细菌具有生物矿化的能力和条件,也无法在不锈钢表面形成矿化膜 (WT实验组),钝化膜的存在能够抑制矿化膜的生成。但一旦钝化膜被具有腐蚀性的黑色素物质破坏,就会为矿化膜的生长创造成核位点,矿化膜不断生长也会继续大面积破坏钝化膜。∆hmgA组的不锈钢样品表面虽然有碳酸钙结构的矿化膜,却无法提供抑制腐蚀的作用,一方面是因为矿化膜的防护效果不如钝化膜好,另一方面也是因为环境中存在具有腐蚀性质的黑色素物质,这两种因素结合在一起,导致产色菌∆hmgA组的不锈钢样品腐蚀的最为严重,表面出现大量的点蚀坑。另外,黑色素溶液中不锈钢样品的钝化膜虽然也遭到破坏,但不锈钢的自钝化功能仍然存在,所以点蚀数量不多。
3 结论
(1) 选用同属同种的P. lipolytica的WT、∆hmgA、黑色素溶液为研究对象,其中,∆hmgA及其黑色素对不锈钢有显著的腐蚀作用,在样品表面出现了明显的点蚀,∆hmgA培养液中的不锈钢样品比黑色素溶液中腐蚀更严重。
(2) P. lipolytica的WT和∆hmgA都能够分泌胞外多糖,具备诱导生物矿化的条件,但只有∆hmgA在不锈钢表面形成生物矿化膜。这说明有钝化膜存在时,不能够形成矿化位点,会阻碍细菌生物矿化;但是当细菌分泌黑色素破坏钝化膜后,就为生物矿化膜的生长提供了条件,生物矿化膜可以不断生长并且替代钝化膜。生物矿化膜的存在使得不锈钢的自钝化能力消失,导致腐蚀加速和点蚀形成。
参考文献
The cost of corrosion in China
[J].
Mechanistic microbiologically influenced corrosion modeling—a review
[J].
At present, most Microbiologically Influenced Corrosion (MIC) models used in the petroleum industry are risk based, and predict the likelihood of MIC. It is imperative that the industry adopts mechanistic models to accurately quantify MIC in pipes/facilities. This paper summarizes the findings of a literature review of mechanistic MIC models available in the public domain. The results indicated that most of the models reviewed consider Sulphate Reducing Bacteria (SRB) as the main players in MIC, and very few models consider biofllm formation and transport phenomena. Furthermore, no models were found to accurately correlate sessile and planktonic bacteria.
Evaluation of ozone for preventing fungal influenced corrosion of reinforced concrete bridges over the River Nile, Egypt
[J].Fungal influenced corrosion (FIC) of some corroded sites in three selected bridges [Embaba bridge (E-bridge), Kasr al-Nile-bridge (K-bridge) and University bridge (U-bridge)] located over the River Nile in Egypt were investigated. Six fungal species, belong to 12 fungal genera, were isolated from the corroded reinforced concrete of the three tested bridges. Fourier transform infrared spectroscopy (FTIR) was screened for the most dominant fungal species (Fusarium oxysporium) which showed in all tested bridges that indicated the presence of amine group accompanied with polysaccharides contents. FIC of the most deteriorated bridge (K-bridge) was documented with FTIR. The association of fungal spores with corrosion products was recorded with scanning electron microscope (SEM). Evaluation of ozone for preventing FIC of the K-bridge was carried out by recording the corrosion rate and the corresponding inhibition efficiency (IE%). No mycelial growth with 100% IE was observed at 3 ppm ozone concentration after 120 min exposure time. With longer duration of ozone exposure, the membrane permeability of F. oxysporium was compromised as indicated by protein and nucleic acid leakages accompanied with lipid and tryptophan oxidation. The total intracellular and extracellular proteins of F. oxysporium were run on sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) indicated the increasing of the supernatant protein on the expense of the cellular protein bands with extending ozone exposure time (0-80 min).
Research progress on microbiological induced corrosion of metallic materials under ocean environment
[J].
海洋环境下金属材料微生物腐蚀研究进展
[J].
Investigation on microbiologically influenced corrosion behavior of CrCoNi medium-entropy alloy by pseudomonas aeruginosa
[J].
铜绿假单胞菌对CrCoNi中熵合金微生物腐蚀行为的影响
[J].采用多种电化学实验手段及场发射扫描电子显微镜(FESEM)、激光共聚焦扫描显微镜(CLSM)等分析技术,结合活死细菌染色实验、点蚀坑深度分析等方法,以316L不锈钢为对比,研究了CrCoNi中熵合金在含铜绿假单胞菌培养基中的微生物腐蚀行为。结果表明:铜绿假单胞菌能够在CrCoNi中熵合金表面形成不均匀的生物被膜,从而降低开路电位,减小极化电阻和电荷转移电阻,增大腐蚀电流密度;铜绿假单胞菌生物被膜在一定程度上破坏了钝化膜,导致浸泡在含铜绿假单胞菌培养基中的CrCoNi中熵合金的最大点蚀坑深度(4.8 μm)大于无菌培养基中CrCoNi中熵合金的最大点蚀坑深度(2.3 μm)。与316L不锈钢相比,CrCoNi中熵合金的开路电位较高,腐蚀电流密度和腐蚀速率较小,钝化膜的修复能力较强,在含铜绿假单胞菌培养基中浸泡后的最大点蚀坑深度小于316L不锈钢(5.8 μm)。
Microbiologically influenced corrosion and mechanisms
[J].
微生物腐蚀及腐蚀机理研究进展
[J].
Effects of biogenic H2S on the microbiologically influenced corrosion of C1018 carbon steel by sulfate reducing Desulfovibrio vulgaris biofilm
[J].
Bacterial pigments and their applications
[J].
Phenazines and other redox-active antibiotics promote microbial mineral reduction
[J].
Melanin production and use as a soluble electron shuttle for Fe(III) oxide reduction and as a terminal electron acceptor by Shewanella algae BrY
[J].
Correlation between pigmentation and antifouling compounds produced by Pseudoalteromonas tunicata
[J].
The corrosion behavior and mechanism of carbon steel induced by extracellular polymeric substances of iron-oxidizing bacteria
[J].
Pitting corrosion behaviour of austenitic stainless steels-combining effects of Mn and Mo additions
[J].
Corrosion and Inhibition of 316L stainless steel in neutral medium by 2-mercaptobenzimidazole
[J].
Electrochemical behaviour of AISI 316 austenitic stainless steel in acidic media containing chloride ions
[J].
Development of an optimized electrochemical process for subsequent coating of 316 stainless steel for stent applications
[J].
Development of an efficient conjugation-based genetic manipulation system for Pseudoalteromonas
[J].
Genome-scale phylogenetic and DNA composition analyses of Antarctic Pseudoalteromonas bacteria reveal inconsistencies in current taxonomic affiliation
[J].
Enhanced corrosion protection action of biofilms based on endogenous and exogenous bacterial cellulose
[J].
Galvanic corrosion behavior of low alloy steel, stainless steel and Al-Mg alloy in simulated deep sea environment
[J].
低合金钢、不锈钢和铝镁合金在模拟深海环境中的电偶腐蚀行为研究
[J].
Corrosion behavior of 316L stainless steel in simulated oilfield wastewater
[J].
S2-对316L不锈钢在模拟油田污水中的腐蚀行为影响研究
[J].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}